Hay cuatro ideas principales en la prevención de recaídas. Primero, la recaída es un proceso gradual con etapas distintas. El objetivo del tratamiento es ayudar a las personas a reconocer las primeras etapas, en las que las posibilidades de éxito son mayores. En segundo lugar, la recuperación es un proceso de crecimiento personal con hitos en el desarrollo. Cada etapa de recuperación tiene sus propios riesgos de recaída. Tercero, las principales herramientas de prevención de recaídas son la terapia cognitiva y la relajación de la mente y el cuerpo, que se utilizan para desarrollar habilidades de afrontamiento saludables. Cuarto, la mayoría de las recaídas pueden explicarse en términos de algunas reglas básicas.
Palabras clave: recaída, prevención de recaídas, cinco reglas de recuperación, etapas de recaída, recaída emocional, recaída mental, recaída física, autocuidado, negación, situaciones de alto riesgo, terapia cognitiva, relajación mente-cuerpo, terapia de prevención de recaídas basada en la atención plena , grupos de autoayuda, grupos de 12 pasos, Alcohólicos Anónimos, Narcóticos Anónimos, etapas de recuperación, etapa de abstinencia, etapa de reparación, etapa de crecimiento, retirada post-aguda, PAWS, no usuario, usuario rechazado.
Introducción
La prevención de recaídas es la razón por la cual la mayoría de las personas buscan tratamiento. Para cuando la mayoría de las personas buscan ayuda, ya han intentado dejar de fumar por su cuenta y están buscando una mejor solución. Este artículo ofrece un enfoque práctico para la prevención de recaídas que funciona bien tanto en terapia individual como grupal.
Hay cuatro ideas principales en la prevención de recaídas. Primero, la recaída es un proceso gradual con etapas distintas. El objetivo del tratamiento es ayudar a las personas a reconocer las primeras etapas, en las que las posibilidades de éxito son mayores [1]. En segundo lugar, la recuperación es un proceso de crecimiento personal con hitos en el desarrollo. Cada etapa de recuperación tiene sus propios riesgos de recaída [2]. En tercer lugar, las principales herramientas de prevención de recaídas son la terapia cognitiva y la relajación cuerpo-mente, que cambian el pensamiento negativo y desarrollan habilidades de afrontamiento saludables [3]. Cuarto, la mayoría de las recaídas pueden explicarse en términos de algunas reglas básicas [4]. Educar a los pacientes en estas pocas reglas puede ayudarlos a enfocarse en lo que es importante.
Me gustaría aprovechar esta oportunidad, después de haber sido invitado a presentar mi perspectiva sobre la prevención de recaídas, para proporcionar una visión general del campo y documentar algunas ideas en medicina para la adicción que son ampliamente aceptadas pero que aún no se han introducido en la literatura. También he incluido un enlace a un video de servicio público sobre prevención de recaídas que contiene muchas de las ideas de este artículo y que está disponible gratuitamente para individuos e instituciones [5].
Las etapas de la recaída
La clave para la prevención de recaídas es comprender que la recaída ocurre gradualmente [6]. Comienza semanas y en algún momento meses antes de que un individuo tome una bebida o droga. El objetivo del tratamiento es ayudar a las personas a reconocer los primeros signos de advertencia de una recaída y desarrollar habilidades de afrontamiento para prevenir la recaída al comienzo del proceso, cuando las posibilidades de éxito son mayores. Se ha demostrado que esto reduce significativamente el riesgo de recaída [7]. Gorski ha dividido la recaída en 11 fases [6]. Este nivel de detalle es útil para los médicos, pero a veces puede ser abrumador para los pacientes. Me ha resultado útil pensar en términos de tres etapas de recaída: emocional, mental y física [4].
Recaída emocional
Durante la recaída emocional, las personas no piensan en consumir. Recuerdan su última recaída y no quieren repetirla. Pero sus emociones y comportamientos los están preparando para una recaída en el camino. Debido a que los pacientes no están pensando conscientemente en consumir durante esta etapa, la negación es una gran parte de la recaída emocional.
Estos son algunos de los signos de recaída emocional [1]: 1) reprimir emociones; 2) aislamiento; 3) no ir a las reuniones; 4) ir a reuniones, pero no compartir; 5) enfocarse en los demás (enfocarse en los problemas de otras personas o enfocarse en cómo otras personas los afectan); y 6) malos hábitos alimenticios y de sueño. El denominador común de la recaída emocional es el mal autocuidado, en el que el autocuidado se define ampliamente para incluir el cuidado emocional, psicológico y físico.
Uno de los objetivos principales de la terapia en esta etapa es ayudar a los pacientes a comprender qué significa el autocuidado y por qué es importante [4]. La necesidad de autocuidado varía de persona a persona. Un simple recordatorio de mal cuidado personal es el acrónimo HALT: hambriento, enojado, solo y cansado (hungry, angry, lonely, and tired). Para algunas personas, el autocuidado es tan básico como el autocuidado físico, como el sueño, la higiene y una dieta saludable. Para la mayoría de las personas, el autocuidado se trata del autocuidado emocional. Los pacientes necesitan darse tiempo para ellos mismos, ser amables con ellos mismos y darse permiso para divertirse. Estos temas generalmente deben revisarse muchas veces durante la terapia: “¿Estás comenzando a sentirte exhausto nuevamente? ¿Sientes que estás siendo bueno tú mismo? Como te diviertes ¿Estás ahorrando tiempo para ti o estás atrapado en la vida?
Otro objetivo de la terapia en esta etapa es ayudar a los pacientes a identificar su negación. Encuentro útil alentar a los pacientes a comparar su comportamiento actual con el comportamiento durante las recaídas pasadas y ver si su autocuidado está empeorando o mejorando.
La transición entre la recaída emocional y mental no es arbitraria, sino la consecuencia natural de un cuidado personal prolongado y deficiente. Cuando las personas exhiben un mal cuidado personal y viven una recaída emocional el tiempo suficiente, eventualmente comienzan a sentirse incómodos en su propia piel. Comienzan a sentirse inquietos, irritables y descontentos. A medida que aumenta su tensión, comienzan a pensar en consumir solo para escapar.
Recaída mental
En la recaída mental, hay una guerra en la mente de las personas. Parte de ellos quiere consumir, pero parte de ellos no. A medida que los individuos profundizan en la recaída mental, su resistencia cognitiva a la recaída disminuye y su necesidad de escapar aumenta.
Estos son algunos de los signos de recaída mental [1]: 1) ansia de drogas o alcohol; 2) pensar en personas, lugares y cosas asociadas con el uso pasado; 3) minimizar las consecuencias del uso pasado o idealizar el consumo pasado; 4) negociación; 5) mentir; 6) pensar en esquemas para controlar mejor el consumo; 7) buscando oportunidades de recaída; y 8) planear una recaída.
Ayudar a los pacientes a evitar situaciones de alto riesgo es un objetivo importante de la terapia. La experiencia clínica ha demostrado que las personas tienen dificultades para identificar sus situaciones de alto riesgo y creer que son de alto riesgo. A veces piensan que evitar situaciones de alto riesgo es un signo de debilidad.
En la negociación, las personas comienzan a pensar en escenarios en los que sería aceptable su uso. Un ejemplo común es cuando las personas se dan permiso para consumir en vacaciones o en un viaje. Es una experiencia común que los aeropuertos y centros turísticos con todo incluido son entornos de alto riesgo en la recuperación temprana. Otra forma de negociación es cuando las personas comienzan a pensar que pueden recaer periódicamente, tal vez de manera controlada, por ejemplo, una o dos veces al año. La negociación también puede tomar la forma de cambiar una sustancia adictiva por otra.
Ocasionales, breves pensamientos de uso son normales en la recuperación temprana y son diferentes de la recaída mental. Cuando las personas ingresan a un programa de abuso de sustancias, a menudo las escucho decir: «No quiero tener que pensar nunca en volver a usarlas». Puede ser aterrador cuando descubren que todavía tienen antojos ocasionales. Sienten que están haciendo algo mal y que se han decepcionado a sí mismos y a sus familias. A veces son reacios incluso a mencionar pensamientos de uso porque están muy avergonzados por ellos.
La experiencia clínica ha demostrado que los pensamientos ocasionales sobre el uso deben normalizarse en la terapia. No significan que el individuo recaerá o que están haciendo un mal trabajo de recuperación. Una vez que una persona ha experimentado adicción, es imposible borrar la memoria. Pero con buenas habilidades de afrontamiento, una persona puede aprender a dejar de consumir rápidamente.
Los médicos pueden distinguir la recaída mental de los pensamientos ocasionales de uso al monitorear el comportamiento de un paciente longitudinalmente. Las señales de advertencia son cuando los pensamientos de consumir cambian en el carácter, se vuelven más insistentes o aumentan en frecuencia.
Recaída física
Finalmente, la recaída física es cuando un individuo comienza a consumir nuevamente. Algunos investigadores dividen la recaída física en un «lapso» (la bebida inicial o el uso de drogas) y una «recaída» (un retorno al uso no controlado) [8]. La experiencia clínica ha demostrado que cuando los clientes se concentran demasiado en cuánto consumieron durante un lapso, no aprecian completamente las consecuencias de una bebida. Una vez que un individuo ha tomado una bebida o un consumo de drogas, puede provocar rápidamente una recaída del uso incontrolado. Pero lo que es más importante, generalmente conducirá a una recaída mental de pensamiento obsesivo o descontrolado sobre el uso, lo que eventualmente puede conducir a una recaída física.
La mayoría de las recaídas físicas son recaídas de oportunidad. Ocurren cuando la persona tiene una ventana en la que siente que no la atraparán. Parte de la prevención de recaídas implica ensayar estas situaciones y desarrollar estrategias de salida saludables.
Cuando las personas no entienden la prevención de recaídas, piensan que implica decir no justo antes de que estén a punto de usar. Pero esa es la etapa final y más difícil de detener, razón por la cual las personas recaen. Si un individuo permanece en una recaída mental el tiempo suficiente sin las habilidades de afrontamiento necesarias, la experiencia clínica ha demostrado que es más probable que recurra a las drogas o al alcohol solo para escapar de su confusión.
Terapia cognitiva y prevención de recaídas
La terapia cognitiva es una de las principales herramientas para cambiar el pensamiento negativo de las personas y desarrollar habilidades de afrontamiento saludables [9, 10]. La efectividad de la terapia cognitiva en la prevención de recaídas ha sido confirmada en numerosos estudios [11].
Esta es una breve lista de los tipos de pensamiento negativo que son obstáculos para la recuperación y son temas para la terapia cognitiva [9]: 1) Mi problema se debe a otras personas; 2) No creo que pueda manejar la vida sin consumir; 3) Tal vez solo pueda usar ocasionalmente; 4) La vida no será divertida, no seré divertido sin consumir; 5) Me preocupa convertirme en alguien que no me gusta; 6) No puedo hacer todos los cambios necesarios; No puedo cambiar a mis amigos; 7) No quiero abandonar a mi familia; 8) La recuperación es demasiado trabajo; 9) Mis antojos serán abrumadores; No podré resistirme a ellos; 10) Si me detengo, solo comenzaré de nuevo; Nunca he terminado nada; 11) Nadie tiene que saber si recaigo; y 12) Me preocupa que mi adicción me haya dañado tanto que no pueda recuperarme.
El pensamiento negativo que subyace al pensamiento adictivo suele ser todo o nada, descalificando lo positivo, y viéndolo todo negativo [9]. Estos pensamientos pueden provocar ansiedad, resentimientos, estrés y depresión, todo lo cual puede conducir a una recaída. La terapia cognitiva y la relajación de la mente y el cuerpo ayudan a romper los viejos hábitos y reentrenar los circuitos neuronales para crear formas de pensar nuevas y más saludables [12, 13].
Temor
El miedo es un patrón de pensamiento negativo común en la adicción [14]. Estas son algunas de las categorías del pensamiento temeroso: 1) miedo a no estar a la altura; 2) miedo a ser juzgado; 3) miedo a sentirse como un fraude y ser descubierto; 4) miedo a no saber vivir en el mundo sin drogas ni alcohol; 5) miedo al éxito; y 6) miedo a la recaída.
Un miedo básico a la recuperación es que el individuo no es capaz de recuperarse. La creencia es que la recuperación requiere una fuerza especial o fuerza de voluntad que el individuo no posee. Las recaídas pasadas se toman como prueba de que el individuo no tiene lo necesario para recuperarse [9]. La terapia cognitiva ayuda a los clientes a ver que la recuperación se basa en las habilidades de afrontamiento y no en la fuerza de voluntad.
Redefiniendo la diversión
Una de las tareas importantes de la terapia es ayudar a las personas a redefinir la diversión. La experiencia clínica ha demostrado que cuando los clientes están estresados, tienden a dar glamour a su consumo anterior y a pensarlo con nostalgia. Empiezan a pensar que la recuperación es un trabajo duro y la adicción fue divertida. Comienzan a descalificar los aspectos positivos que han obtenido a través de la recuperación. El desafío cognitivo es reconocer que la recuperación es a veces un trabajo duro, pero la adicción es aún más difícil. Si la adicción fuera tan fácil, las personas no querrían dejar de fumar y no tendrían que dejar de hacerlo.
Cuando las personas continúan refiriéndose a sus días de consumo como «diversión», continúan minimizando las consecuencias negativas de la adicción. La teoría de la expectativa ha demostrado que cuando las personas esperan divertirse, generalmente lo hacen, y cuando esperan que algo no sea divertido, generalmente no lo es [15] En las primeras etapas del abuso de sustancias, el uso es principalmente una experiencia positiva para aquellos que están predispuestos emocional y genéticamente. Más tarde, cuando el uso se convierte en una experiencia negativa, a menudo continúan esperando que sea positivo. Es común escuchar a los adictos hablar sobre perseguir los primeros momentos que tenían. Por otro lado, las personas esperan que no usar drogas o alcohol les provoque el dolor emocional o el aburrimiento del que trataron de escapar. Por lo tanto, por un lado, las personas esperan que el uso continúe siendo divertido y, por otro lado, esperan que no usar sea incómodo. La terapia cognitiva puede ayudar a abordar estos dos conceptos erróneos.
Aprendiendo de los contratiempos
La forma en que las personas enfrentan los contratiempos juega un papel importante en la recuperación. Un revés puede ser cualquier comportamiento que acerque a un individuo a una recaída física. Algunos ejemplos de contratiempos no son establecer límites saludables, no pedir ayuda, no evitar situaciones de alto riesgo y no practicar el cuidado personal. Un revés no tiene que terminar en una recaída para ser digno de discusión en la terapia.
Las personas en recuperación tienden a ver los reveses como fracasos porque son inusualmente duros con ellos mismos [9]. Los contratiempos pueden establecer un círculo vicioso, en el que las personas ven los contratiempos como una confirmación de su visión negativa de sí mismos. Sienten que no pueden vivir la vida en los términos de la vida. Esto puede conducir a un mayor uso y una mayor sensación de fracaso. Finalmente, dejan de centrarse en el progreso que han logrado y comienzan a ver el camino por delante como abrumador [16].
Los contratiempos son una parte normal del progreso. No son fracasos. Son causadas por habilidades de afrontamiento insuficientes y / o planificación inadecuada, que son problemas que pueden solucionarse [8]. Se alienta a los pacientes a desafiar su pensamiento al observar los éxitos pasados y reconocer las fortalezas que aportan a la recuperación [8]. Esto impide que los pacientes hagan declaraciones globales, como «Esto prueba que soy un fracaso«. Cuando las personas adoptan una visión dicotómica de recuperación de todo o nada, es más probable que se sientan abrumados y abandonen los objetivos a largo plazo en favor del alivio a corto plazo. Esta reacción se denomina efecto de violación de la abstinencia [8].
Cómo sentirse cómodo con estar incómodo
En términos más generales, creo que las personas en recuperación necesitan aprender a sentirse cómodas con sentirse incómodas. A menudo asumen que los no adictos no tienen los mismos problemas o experimentan las mismas emociones negativas. Por lo tanto, sienten que es defendible o necesario escapar de sus sentimientos negativos. El desafío cognitivo es indicar que los sentimientos negativos no son signos de fracaso, sino una parte normal de la vida y oportunidades de crecimiento. Ayudar a los pacientes a sentirse cómodos con la incomodidad puede reducir su necesidad de escapar hacia la adicción.
Las etapas de recuperación
La recuperación es un proceso de crecimiento personal en el que cada etapa tiene sus propios riesgos de recaída y sus propias tareas de desarrollo para llegar a la siguiente etapa [2]. Las etapas de recuperación no son del mismo largo para cada persona, pero son una forma útil de ver la recuperación y enseñar la recuperación a los pacientes. En términos generales, hay tres etapas de recuperación. En el modelo de desarrollo original, las etapas se llamaban «transición, recuperación temprana y recuperación continua» [2]. Los nombres más descriptivos pueden ser «abstinencia, reparación y crecimiento».
Etapa de abstinencia
Se sostiene comúnmente que la etapa de abstinencia comienza inmediatamente después de que una persona deja de usar y generalmente dura de 1 a 2 años [1]. El enfoque principal de esta etapa es lidiar con los antojos (craving) y no consumir. Estas son algunas de las tareas de la etapa de abstinencia [2]:
Acepta que tienes una adicción
Practica la honestidad en la vida
Desarrolla habilidades de afrontamiento para lidiar con los antojos
Participa activamente en grupos de autoayuda
Practica el cuidado personal y decir no
Comprende las etapas de la recaída.
Deshágase de los amigos que están consumiendo
Comprende los peligros de la adicción cruzada
Trata con la abstinencia post aguda
Desarrolla alternativas saludables al consumo
Mírate a ti mismo como un no consumidor
Hay muchos riesgos para la recuperación en esta etapa, incluidos los antojos físicos, el mal cuidado personal, querer consumir solo una vez más y luchar para saber si uno tiene una adicción. Los pacientes a menudo están ansiosos por hacer grandes cambios externos en la recuperación temprana, como cambiar de trabajo o terminar una relación. En general, se considera que deben evitarse los grandes cambios en el primer año hasta que las personas tengan suficiente perspectiva para ver su papel, si lo hay, en estos temas y no centrarse por completo en los demás.
Las tareas de esta etapa se pueden resumir en una mejora del autocuidado físico y emocional. La experiencia clínica ha demostrado que las personas en recuperación a menudo tienen prisa por saltarse estas tareas y continuar con lo que creen que son los verdaderos problemas de la recuperación. Se debe recordar a los pacientes que la falta de autocuidado es lo que los trajo aquí y que la continua falta de autocuidado conducirá a una recaída.
Abstinencia post-aguda
Lidiar con la abstinencia post aguda es una de las tareas de la etapa de abstinencia [1]. La abstinencia post aguda comienza poco después de la fase aguda de abstinencia y es una causa común de recaída [17]. A diferencia de la abstinencia aguda, que tiene principalmente síntomas físicos, el síndrome de abstinencia post-aguda (PAWS) tiene principalmente síntomas psicológicos y emocionales. Sus síntomas también tienden a ser similares para la mayoría de las adicciones, a diferencia de la abstinencia aguda, que tiende a tener síntomas específicos para cada adicción [1].
Estos son algunos de los síntomas de la abstinencia post-aguda [1, 18, 19]: 1) cambios de humor; 2) ansiedad; 3) irritabilidad; 4) energía variable; 5) poco entusiasmo; 6) concentración variable; y 7) sueño perturbado. Muchos de los síntomas de abstinencia post-aguda se superponen con la depresión, pero se espera que los síntomas de abstinencia post-aguda mejoren gradualmente con el tiempo [1].
Probablemente, lo más importante que hay que entender sobre la abstinencia post-aguda es su duración prolongada, que puede durar hasta 2 años [1, 20]. El peligro es que los síntomas tienden a ir y venir. No es inusual no tener síntomas durante 1 a 2 semanas, solo para ser golpeado nuevamente [1]. Este momento es cuando las personas corren el riesgo de recaída, cuando no están preparados para la naturaleza prolongada de la abstinencia post-aguda. La experiencia clínica ha demostrado que cuando los pacientes luchan con la abstinencia post-aguda, tienden a ver muy negras sus posibilidades de recuperación. Piensan que no están progresando. El desafío cognitivo es alentar a los clientes a medir su progreso mes a mes en lugar de día a día o semana a semana.
Etapa de reparación
En la segunda etapa de recuperación, la tarea principal es reparar el daño causado por la adicción [2]. La experiencia clínica ha demostrado que esta etapa generalmente dura de 2 a 3 años.
En la etapa de abstinencia de recuperación, los pacientes generalmente se sienten cada vez mejor. Finalmente están tomando el control de sus vidas. Pero en la etapa de reparación de la recuperación, no es inusual que las personas se sientan peor durante un tiempo. Deben enfrentar el daño causado por la adicción a sus relaciones, empleo, finanzas y autoestima. También deben superar la culpa y el autoetiquetado negativo que evolucionó durante la adicción. Los pacientes a veces piensan que han sido tan dañados por su adicción que no pueden experimentar alegría, sentirse seguros o tener relaciones saludables [9].
Estas son algunas de las tareas de desarrollo de la etapa de reparación de recuperación [1, 2]:
Usa la terapia cognitiva para superar el autoetiquetado negativo y la catástrofe
Comprende que los individuos no son tu adicción.
Repara las relaciones y haz las paces cuando sea posible
Comienza a sentirte cómodo con la incomodidad
Mejora el autocuidado y conviértelo en una parte integral de la recuperación
Desarrolla un estilo de vida equilibrado y saludable.
Continúa participando en grupos de autoayuda
Desarrolla alternativas más saludables que el consumo
La experiencia clínica ha demostrado que las causas comunes de recaída en esta etapa son los malos cuidados personales y no ir a grupos de autoayuda.
Etapa de crecimiento
La etapa de crecimiento consiste en desarrollar habilidades que las personas tal vez nunca hayan aprendido y que las predispongan a la adicción [1, 2]. La etapa de reparación de la recuperación se trataba de ponerse al día, y la etapa de crecimiento se trata de avanzar. La experiencia clínica ha demostrado que esta etapa generalmente comienza de 3 a 5 años después de que las personas han dejado de consumir drogas o alcohol y es un camino de por vida.
Este es también el momento de tratar cualquier problema de la familia de origen o cualquier trauma pasado que pueda haber ocurrido. Estos son problemas que los pacientes a veces están ansiosos por resolver. Pero pueden ser problemas estresantes y, si se abordan demasiado pronto, los pacientes pueden no tener las habilidades de afrontamiento necesarias para manejarlos, lo que puede conducir a una recaída.
Estas son algunas de las tareas de la etapa de crecimiento [1, 2]:
Identifica y repara el pensamiento negativo y los patrones autodestructivos.
Comprende cómo se han transmitido los patrones familiares negativos, lo que ayudará a las personas a dejar de lado los resentimientos y seguir adelante
Desafía los miedos con la terapia cognitiva y la relajación de la mente y el cuerpo.
Establece límites saludables
Comienza a retribuir y ayudar a otros
Reevalúa periódicamente el estilo de vida y asegúrate de que estás en el camino
Las tareas de esta etapa son similares a las tareas que enfrentan los no adictos en la vida cotidiana. Cuando los no adictos no desarrollan habilidades saludables para la vida, la consecuencia es que pueden ser infelices en la vida. Cuando las personas en recuperación no desarrollan habilidades para la vida saludable, la consecuencia es que también pueden ser infelices en la vida, pero eso puede conducir a una recaída.
Causas de recaída en la etapa tardía de recuperación
En la recuperación de la etapa tardía, las personas están sujetas a riesgos especiales de recaída que a menudo no se ven en las primeras etapas. La experiencia clínica ha demostrado que las siguientes son algunas de las causas de recaída en la etapa de crecimiento de la recuperación.
Los pacientes a menudo quieren dejar atrás su adicción y olvidar que alguna vez tuvieron una adicción. Sienten que han perdido parte de su vida por la adicción y no quieren pasar el resto de su vida enfocados en la recuperación. Empiezan a ir a menos reuniones.
A medida que la vida mejora, las personas comienzan a centrarse menos en el cuidado personal. Asumen más responsabilidades e intentan recuperar el tiempo perdido. En cierto sentido, están tratando de volver a su antigua vida sin el consumo. Dejan de hacer las cosas saludables que contribuyeron a su recuperación.
Los pacientes sienten que no están aprendiendo nada nuevo en las reuniones de autoayuda y comienzan a ir con menos frecuencia. Los pacientes deben comprender que uno de los beneficios de ir a las reuniones es recordar cómo suena la «voz de la adicción«, porque es fácil de olvidar.
Las personas sienten que deberían estar más allá de lo básico. Piensan que es casi vergonzoso hablar sobre los conceptos básicos de la recuperación. Les da vergüenza mencionar que todavía tienen antojos ocasionales o que ya no están seguros de tener una adicción.
Las personas piensan que tienen una mejor comprensión de las drogas y el alcohol y, por lo tanto, piensan que deberían poder controlar una recaída o evitar las consecuencias negativas.
Las cinco reglas de recuperación
Esta sección se basa en mi experiencia de trabajar con adictos durante más de 30 años en programas de tratamiento y en la práctica privada. La experiencia ha demostrado que la mayoría de las recaídas pueden explicarse en términos de algunas reglas básicas [4]. Enseñar a los pacientes estas reglas simples les ayuda a comprender que la recuperación no es complicada ni está fuera de su control. Se basa en unas pocas reglas simples que son fáciles de recordar: 1) cambia tu vida; 2) sé completamente honesto; 3) pide ayuda; 4) practica el autocuidado; y 5) no rompas las reglas.
Regla 1: Cambia tu vida
La regla más importante de recuperación es que una persona no logra la recuperación simplemente por no consumir. La recuperación implica crear una nueva vida en la que es más fácil no consumir. Cuando los individuos no cambian sus vidas, todos los factores que contribuyeron a su adicción eventualmente los alcanzarán.
Pero los pacientes y las familias a menudo comienzan la recuperación esperando no tener que cambiar. A menudo ingresan al tratamiento diciendo: «Queremos recuperar nuestra antigua vida, sin el consumo«. Intento ayudar a los pacientes a comprender que desear recuperar su antigua vida es como desear una recaída. En lugar de ver la necesidad de cambio como algo negativo, se les alienta a ver la recuperación como una oportunidad para el cambio. Si hacen los cambios necesarios, pueden seguir adelante y ser más felices que antes. Este es el «lado positivo» de tener una adicción. Obliga a las personas a reevaluar sus vidas y hacer cambios que los no adictos no tienen que hacer.
Las personas en recuperación a menudo se sienten abrumadas por la idea del cambio. Como parte de su pensamiento de todo o nada, suponen que el cambio significa que deben cambiar todo en sus vidas. Les ayuda a saber que, por lo general, solo hay que cambiar un pequeño porcentaje de sus vidas. También puede ser seguro saber que la mayoría de las personas tienen los mismos problemas y necesitan hacer cambios similares.
Ejemplos de cambio
¿Qué necesita cambiar la mayoría de las personas? Hay tres categorías:
Cambiar los patrones de pensamiento negativo discutidos anteriormente
Evitar personas, lugares y cosas asociadas con el consumo
Incorporar las cinco reglas de recuperación.
Los pacientes deben desarrollar un miedo saludable a las personas, los lugares y las cosas que formaron parte del consumo. Pero esto requiere un entrenamiento mental significativo porque esas personas, lugares y cosas se asociaron previamente con emociones positivas. Además, los pacientes tienden a pensar que desarrollar un miedo saludable a estas cosas es mostrar debilidad o aceptar la derrota.
Regla 2: Sé completamente honesto
La adicción requiere mentir. Los adictos deben mentir acerca de obtener su droga, esconderla, negar las consecuencias y planificar su próxima recaída. Finalmente, los individuos adictos terminan mintiéndose a sí mismos. La experiencia clínica muestra que cuando los pacientes sienten que no pueden ser completamente honestos, es una señal de recaída emocional. A menudo se dice que las personas en recuperación están tan enfermas como sus secretos. Uno de los desafíos de la terapia es ayudar a los pacientes a practicar decir la verdad y a admitir cuando han dicho mal y corregirlo rápidamente.
¿Qué tan honesto debe ser una persona sin poner en peligro su trabajo o sus relaciones? Se alienta a los clientes a comprender el concepto de un círculo de recuperación. Este es un grupo de personas que incluye familiares, médicos, consejeros, grupos de autoayuda y padrinos. Se alienta a las personas a ser completamente honestas dentro de su círculo de recuperación. A medida que los clientes se sientan más cómodos, pueden elegir expandir el tamaño de su círculo.
Probablemente, la interpretación errónea más común de la honestidad completa es cuando los individuos sienten que deben ser honestos acerca de lo que está mal con otras personas. La honestidad, por supuesto, es honestidad. Me gusta decirles a los pacientes que una prueba simple de honestidad completa es que deben sentirse «incómodamente honestos» cuando comparten dentro de su círculo de recuperación. Esto es especialmente importante en los grupos de autoayuda en los que, después de un tiempo, las personas a veces comienzan a realizar los movimientos de participación.
Una pregunta común sobre la honestidad es qué tan honesta debe ser una persona cuando se trata de mentiras pasadas. La respuesta general es que la honestidad siempre es preferible, excepto cuando puede dañar a otros [14, 21].
Regla 3: pedir ayuda
La mayoría de las personas comienzan la recuperación tratando de hacerlo por su cuenta. Quieren demostrar que tienen control sobre su adicción y que no son tan poco saludables como la gente piensa. Se ha demostrado que unirse a un grupo de autoayuda aumenta significativamente las posibilidades de recuperación a largo plazo. La combinación de un programa de abuso de sustancias y un grupo de autoayuda es la más efectiva [22, 23].
Hay muchos grupos de autoayuda para elegir. Los grupos de doce pasos incluyen Alcohólicos Anónimos (AA), Narcóticos Anónimos (NA), Marihuana Anónimos (MA), Cocaína Anónima (CA), Jugadores Anónimos (GA) e Hijos Adultos de Alcohólicos (ACA). Cada país, cada ciudad y casi todos los cruceros tienen una reunión de 12 pasos. Hay otros grupos de autoayuda, incluidos Mujeres para la sobriedad, Organizaciones seculares para la sobriedad, Recuperación inteligente y grupos Caduceus para profesionales de la salud. Se ha demostrado que la forma de aprovechar al máximo los grupos de 12 pasos es asistir a reuniones regularmente, tener un padrino, leer materiales de 12 pasos y tener un objetivo de abstinencia [24, 25].
Estos son algunos de los beneficios generalmente reconocidos de la participación activa en grupos de autoayuda: 1) las personas sienten que no están solas; 2) aprenden cómo suena la voz de la adicción al escucharla en otros; 3) aprenden cómo otras personas se han recuperado y qué habilidades de afrontamiento han tenido éxito; y 4) tienen un lugar seguro para ir donde no serán juzgados.
Hay un beneficio de los grupos de autoayuda que merece atención especial. La culpa y la vergüenza son emociones comunes en la adicción [26]. Pueden ser obstáculos para la recuperación, porque las personas pueden sentir que han sido dañadas por su adicción y no merecen la recuperación o la felicidad. La experiencia clínica ha demostrado que los grupos de autoayuda ayudan a las personas a superar su culpa y vergüenza de la adicción al ver que no están solos. Sienten que la recuperación está a su alcance.
Estas son algunas de las razones que dan los clientes por no unirse a grupos de autoayuda: 1) Si me uno a un grupo, estaría admitiendo que soy adicto o alcohólico; 2) Quiero hacerlo por mi cuenta; 3) No me gustan los grupos; 4) No soy un ensamblador; 5) No me gusta hablar frente a otras personas; 6) No quiero cambiar de una adicción a volverme adicta a AA; 7) Me temo que seré reconocido; y 8) No me gustan los aspectos religiosos. El pensamiento negativo en todas estas objeciones es material para la terapia cognitiva.
Regla 4: Practica el autocuidado
Para comprender la importancia del autocuidado, es útil entender por qué la mayoría de las personas usan drogas y alcohol. La mayoría de las personas usan para escapar, relajarse o recompensarse [4]. Estos son los principales beneficios del uso. Ayuda a reconocer estos beneficios en la terapia para que las personas puedan comprender la importancia del autocuidado y estar motivados para encontrar alternativas saludables.
A pesar de su importancia, el autocuidado es uno de los aspectos más olvidados de la recuperación. Sin ella, las personas pueden ir a reuniones de autoayuda, tener un padrino, hacer un trabajo paso a paso y aun así recaer. El cuidado personal es difícil porque las personas en recuperación tienden a ser difíciles para sí mismas [9]. Esto puede presentarse abiertamente, como individuos que no sienten que merecen ser buenos consigo mismos o que tienden a ser los últimos, o puede aparecer encubiertamente como individuos que dicen que pueden ser buenos consigo mismos pero que en realidad son despiadadamente críticos de sí mismos. El cuidado personal es especialmente difícil para los hijos adultos de adictos [27].
Una pieza faltante del rompecabezas para muchos clientes es comprender la diferencia entre el egoísmo y el cuidado personal. El egoísmo es tomar más de lo que una persona necesita. El cuidado personal es tomar todo lo que uno necesita. La experiencia clínica ha demostrado que las personas adictas generalmente toman menos de lo que necesitan y, como resultado, se agotan o se resienten y recurren a su adicción para relajarse o escapar. Parte del pensamiento adictivo desafiante es alentar a los clientes a ver que no pueden ser buenos con los demás si primero no son buenos con ellos mismos.
Las personas usan drogas y alcohol para escapar de las emociones negativas; sin embargo, también se usan como recompensa y / o para mejorar las emociones positivas [11]. El mal cuidado personal también juega un papel en estas situaciones. En estas situaciones, el mal cuidado personal a menudo precede al consumo de drogas o alcohol. Por ejemplo, las personas trabajan duro para lograr un objetivo, y cuando se logra, quieren celebrar. Pero como parte de su pensamiento de todo o nada, mientras trabajaban, sintieron que no merecían una recompensa hasta que el trabajo estuviera terminado. Como no se permitieron pequeñas recompensas durante el trabajo, la única recompensa que será suficiente al final es una gran recompensa, que en el pasado significaba consumir.
Autocuidado: Relajación cuerpo-mente
Numerosos estudios han demostrado que la relajación cuerpo-mente reduce el uso de drogas y alcohol y es eficaz en la prevención de recaídas a largo plazo [28, 29]. La terapia de prevención de recaídas y la relajación mente-cuerpo se combinan comúnmente en la prevención de recaídas basada en el mindfulness [30].
La relajación cuerpo-mente juega varios roles en la recuperación [4]. Primero, el estrés y la tensión son desencadenantes comunes de la recaída. En segundo lugar, la relajación de la mente y el cuerpo ayuda a las personas a abandonar el pensamiento negativo, como detenerse en el pasado o preocuparse por el futuro, que son los desencadenantes de una recaída. Tercero, la relajación mente-cuerpo es una forma de ser amable con uno mismo. La práctica del autocuidado durante la relajación cuerpo-mente se traduce en autocuidado en el resto de la vida. Parte de crear una nueva vida en recuperación es encontrar tiempo para relajarse.
Regla 5: no romper las reglas
El propósito de esta regla es recordar a las personas que no se resistan o saboteen el cambio al insistir en que se recuperen a su manera. Una prueba simple de si una persona está rompiendo las reglas es si busca lagunas en la recuperación. Una señal de advertencia es cuando los pacientes solicitan ayuda profesional e ignoran constantemente el consejo.
En términos generales, una vez que los pacientes han estado en recuperación durante un tiempo, se pueden dividir en dos categorías: no usuarios y usuarios denegados. Los no usuarios dicen que usar fue divertido, pero reconocen que últimamente no lo ha sido. Quieren comenzar el próximo capítulo de su vida.
Los usuarios denegados no quieren o no pueden reconocer completamente el alcance de su adicción. No pueden imaginar la vida sin consumir. Los usuarios denegados invariablemente hacen un trato secreto con ellos mismos que en algún momento intentarán usar nuevamente. Los hitos importantes, como los aniversarios de recuperación, a menudo se consideran razones para consumir. Alternativamente, una vez que se alcanza un hito, las personas sienten que se han recuperado lo suficiente como para poder determinar cuándo y cómo consumir de manera segura. Es notable cuántas personas han recaído de esta manera 5, 10 o 15 años después de la recuperación.
Se alienta a los pacientes a identificar si son no usuarios o usuarios denegados. Un usuario denegado está en una recaída mental crónica y en alto riesgo de una recaída futura. La experiencia clínica ha demostrado que todos los que se encuentran en recuperación temprana son usuarios denegados. El objetivo es ayudar a las personas a pasar de usuarios denegados a no usuarios.
Resumen y conclusiones
Las personas no logran la recuperación simplemente por no consumir. La recuperación implica crear una nueva vida en la que es más fácil no consumir. Si las personas no cambian sus vidas, entonces todos los factores que contribuyeron a su adicción seguirán ahí. Pero la mayoría de las personas comienzan la recuperación con la esperanza de recuperar su antigua vida sin el consumo. La recaída es un proceso gradual que comienza semanas y, a veces, meses antes de que un individuo tome una bebida o droga. Hay tres etapas para la recaída: emocional, mental y física. El denominador común de la recaída emocional es el pobre autocuidado. Si las personas no practican el cuidado personal suficiente, pronto comenzarán a sentirse incómodos en su propia piel y buscarán formas de escapar, relajarse o recompensarse. El objetivo del tratamiento es ayudar a las personas a reconocer los primeros signos de advertencia de recaída y desarrollar habilidades de afrontamiento para prevenir una recaída temprana, cuando las posibilidades de éxito son mayores. La mayoría de las recaídas pueden explicarse en términos de algunas reglas básicas. Comprender estas reglas puede ayudar a los clientes a centrarse en lo que es importante: 1) cambiar su vida; 2) ser completamente honesto; 3) pedir ayuda; 4) practicar el autocuidado; y 5) no romper las reglas.
Referencias
Gorski T, Miller M. Staying Sober: A Guide for Relapse Prevention. Independence, MO: Independence Press; 1986.
Brown S. Treating the Alcoholic: A Developmental Model of Recovery. New York: Wiley; 1985.
Marlatt GA, George WH. Relapse prevention: introduction and overview of the model. Br J Addict. 1984;79(3):261–273.
Melemis SM. I Want to Change My Life: How to Overcome Anxiety, Depression and Addiction. Toronto: Modern Therapies; 2010.
Gorski TT, Miller M. Counseling for Relapse Prevention. Independence, MO: Herald House/Independence Press; 1982.
Bennett GA, Withers J, Thomas PW, Higgins DS, Bailey J, Parry L. et al. A randomised trial of early warning signs relapse prevention training in the treatment of alcohol dependence. Addict Behav. 2005;30(6):1111–1124.
Alcoholics Anonymous World Services. Alcoholics Anonymous Big Book. 4th ed. New York: Alcoholics Anonymous World Services; 2001.
Hasking P, Lyvers M, Carlopio C. The relationship between coping strategies, alcohol expectancies, drinking motives and drinking behaviour. Addict Behav. 2011;36(5):479–487.
Tate P. Alcohol: How to Give It Up and Be Glad You Did, A Sensible Approach. 1st ed. Altamonte Springs, FL: Rational Self-Help Press; 1993. [Google Scholar]
Miller WR, Harris RJ. A simple scale of Gorski’s warning signs for relapse. J Stud Alcohol. 2000;61(5):759–765.
Le Bon O, Murphy JR, Staner L, Hoffmann G, Kormoss N, Kentos M. et al. Double-blind, placebo-controlled study of the efficacy of trazodone in alcohol post-withdrawal syndrome: polysomnographic and clinical evaluations. J Clin Psychopharmacol. 2003;23(4):377–383.
Ashton H. In: Comprehensive handbook of drug and alcohol addiction. Miller NS, editor. New York: Dekker; 1991. Protracted Withdrawal Syndromes for Benzodiazepines.
Begleiter H. Brain dysfunction and alcoholism: problems and prospects. Alcohol Clin Exp Res. 1981;5(2):264–266.
Corley MD, Schneider JP. Disclosing Secrets: When, to Whom, & How Much to Reveal. Carefree, AZ: Gentle Path Press; 2002.
Kelly JF, Stout R, Zywiak W, Schneider R. A 3-year study of addiction mutual-help group participation following intensive outpatient treatment. Alcohol Clin Exp Res. 2006;30(8):1381–1392.
Johnson JE, Finney JW, Moos RH. End-of-treatment outcomes in cognitive-behavioral treatment and 12-step substance use treatment programs: do they differ and do they predict 1-year outcomes? J Subst Abuse Treat. 2006;31(1):41–50.
Bradshaw J. Healing the Shame That Binds You. Deerfield Beach, FL: Health Communications; 1988.
Woititz JG. The Complete ACOA Sourcebook: Adult Children of Alcoholics at Home, at Work, and in Love. Deerfield Beach, FL: Health Communications; 2002.
Shafil M, Lavely R, Jaffe R. Meditation and the prevention of alcohol abuse. Am J Psychiatry. 1975;132(9):942–945.
Haré una revisión sobre el concepto de craving o deseo de
consumo y su importancia en las adicciones, proponiendo como líneas de
tratamiento más eficaces los psicofármacos junto con la terapia de corte
cognitivo-conductual. Podría proponer como trabajo describir y analizar la
intervención que se lleva a cabo en Spiral comparándolo con otros modelos de
intervención. Este trabajo se realizó el curso pasado y, por este motivo, se ha
considerado más oportuno abordar el craving, ya que, se considera un fenómeno
básico de los procesos adictivos. En este sentido, para comprender la
naturaleza misma de una adicción, y los diferentes aspectos que ésta engloba,
como son las recaídas, el coste de la abstinencia y la tolerancia, es necesario
conocer el concepto de craving.
En los últimos años el interés por el papel del craving en
las adicciones ha crecido exponencialmente. Su estudio se ha abordado desde muy
distintas perspectivas, dando lugar a diferentes modelos, más aún, en todos
estos modelos el constructo del craving ocupa un papel esencial.
La mayoría de los pacientes en tratamiento por problemas de
adicción hacen mucho hincapié en esta problemática. Continuamente asaltan sobre
su conciencia recuerdos de situaciones de consumo, los cuales, suscitan y
alimentan el deseo por consumir. En un gran número de pacientes en tratamiento
con problemas de drogodependencia, se observa que de modo ininterrumpido notan
un impulso por consumir mucho después que la desintoxicación física ya haya
tenido lugar. En este sentido, el problema de las adicciones, sobre todo de las
adicciones actuales, no es tanto la dependencia física que desaparece
relativamente pronto, como la dependencia psicológica, que en su mayor parte se
explica por aquello en que hemos venido a llamar craving.
Aunque, en los primeros meses de tratamiento la intensidad
del craving es mayor, éste persiste durante meses o incluso años a pesar de
estar abstinente. El deseo irrefrenable por consumir aquella sustancia a la que
se fue adicto, reaparece con cierta frecuencia en sujetos que superaron su
cuadro adictivo de modo exitoso. Es decir, el craving es causa de grandes
recaídas aún después de haber consolidado un estilo de vida saludable y alejado
de cualquier tipo de relación con sustancias adictivas.
En definitiva, el abordaje del craving es de vital importancia en el área de las adicciones, puesto que es uno de los elementos más importantes de recaídas y abandonos terapéuticos.
Las personas que sufren trastornos por el uso de sustancias
desarrollan fuertes asociaciones entre el efecto estimulante de la droga y los estímulos
ambientales que actúan como recordatorios de la experiencia, lo que puede
llevar a una recaída.
Un estudio reciente ha propuesto un mecanismo detrás de la
búsqueda de drogas, la adicción y las recaídas relacionadas con la epigenética.
La epigenética se puede describir como las circunstancias que ocurren en el
entorno que causan que ciertos genes, las instrucciones compuestas de ADN que
controlan las células dentro de nuestro cuerpo, se enciendan o se apaguen.
El Dr. Makoto Taniguchi y Christopher Cowan, de la
Universidad de Medicina de Carolina del Sur y la Escuela de Medicina de
Harvard, descubrieron que la cocaína bloquea un regulador genético particular, cuya
tarea es entrar en las células y evitar que refuercen la asociación entre la
experiencia gratificante de tomar drogas y los factores desencadenantes del
entorno. Como resultado, si el regulador genético no tiene control sobre el
proceso en que se mantiene el enlace entre el entorno y la droga, la memoria y
las señales continúan reforzándose y fortaleciéndose.
Los investigadores creen que este mecanismo puede ser
instrumental en todo el aprendizaje involucrando la vinculación de experiencias
de recompensa de drogas y sus circunstancias asociadas en la memoria. Al
comprender cómo drogas, como la cocaína, influyen en nuestros mecanismos
biológicos, podemos utilizar la información para guiar el desarrollo de un
nuevo tratamiento para los trastornos por uso de sustancias.
El estudio, patrocinado por el NIDA, arroja luz sobre cómo la cocaína crea las poderosas asociaciones de drogas y medio ambiente que subyacen en la búsqueda y recaída de drogas inducidas por estímulos. Los investigadores descubrieron que la cocaína bloquea un proceso epigenético que limita la fuerza de las sinapsis que vinculan experiencias gratificantes con entornos asociados. Los nuevos hallazgos sugieren que un regulador genético llamado histona desacetilasa 5 (HDAC5) puede ofrecer nuevas oportunidades para reducir el riesgo de recaída.
1 Departments of Neuroscience and Psychiatry & Behavioral Sciences, Medical University of South Carolina, Charleston, SC 29425, USA 2 Department of Psychiatry, Harvard Medical School, McLean Hospital, Belmont, MA 02478, USA 3 Neuroscience Graduate Program, University of Texas Southwestern Medical Center, Dallas, TX 75390, USA 4 Department of Brain and Cognitive Science, Massachusetts Institute of Technology, Cambridge, MA 02139, USA 5 Department of Neuroscience, University of Texas Southwestern Medical Center, Dallas, TX 75390, USA 6 Department of Psychiatry, University of Texas Southwestern Medical Center, Dallas, TX 75390, USA 7 Howard Hughes Medical Institute, University of Texas Southwestern Medical Center, Dallas, TX 75390, USA § Lead Contact: ude.csum@cnawoc, Tel: (843) 792-2935 8 Current address: Center of Neuroscience, Institute for Scientific Research and High Technology Services of Panama (INDICASAT AIP), Panamá, Republic of Panamá 9 Current address: Department of Neuroscience, University of Minnesota, Minneapolis, MN 55455, USA 10 Current address: Department of Physiology and Systems Bioscience, Kyoto Prefectural University of Medicine, Kyoto 602-8566, Japan 11 Current address: Department of Neuroscience and Experimental Therapeutics, Texas A&M Health Science Center, Bryan, TX 77807, USA * These authors contributed equally
La recaída es un proceso, no es un evento. Para comprender la prevención de recaídas, debe comprender las etapas de la recaída. La recaída comienza semanas o incluso meses antes del evento de recaída física. En esta página, aprenderá los signos de alerta temprana de recaída y las técnicas específicas de prevención de recaídas para cada etapa de recaída. Hay tres etapas de recaída. (1)
Recaída emocional
Recaída mental
Recaída física
Recaída emocional
En la recaída emocional, no está pensando en tomar. Pero sus emociones y conductas lo están preparando para una posible recaída en el futuro.
Los signos de recaída emocional son:
Ansiedad
Intolerancia
Enfado
La defensa
Cambios de humor
Aislamiento
No pedir ayuda
No ir a reuniones
Los malos hábitos alimenticios
Malos hábitos de sueño
Los signos de recaída emocional son también los síntomas de la abstinencia post-aguda. Si entiende que la abstinencia aguda es más fácil evitar la recaída, porque es más fácil retirarse de la etapa temprana de la recaída. En las etapas posteriores, la fuerza de la recaída se hace más fuerte y la secuencia de eventos se mueve más rápido.
Prevención de la recaída temprana
La prevención de recaídas en esta etapa significa reconocer que estás en recaída emocional y cambiar tu comportamiento. Reconozca que está aislándose y recuerde pedir ayuda. Reconoce que estás ansioso y practica técnicas de relajación. Reconozca que sus hábitos de sueño y alimentación están disminuyendo y practique el cuidado personal.
Si no cambia su comportamiento en esta etapa y vive demasiado tiempo en la etapa de recaída emocional, se agotará, y cuando esté agotado querrá escapar, lo que lo llevará a una recaída mental.
Practica el autocuidado. Lo más importante que puede hacer para prevenir una recaída en esta etapa es cuidarse mejor. Piensa en por qué lo usas. Usted usa drogas o alcohol para escapar, relajarse o recompensarse. Por lo tanto, recae cuando no se cuida y crea situaciones que drenan mental y emocionalmente que le dan ganas de escapar.
Por ejemplo, si no se cuida y come mal o tiene malos hábitos de sueño, se sentirá agotado y querrá escapar. Si no dejas ir tus resentimientos y temores a través de alguna forma de relajación, aumentarán hasta el punto en que te sentirás incómodo en tu propia piel. Si no pides ayuda, te sentirás aislado. Si alguna de esas situaciones continúa por mucho tiempo, comenzará a pensar en usar. Pero si practica el autocuidado, puede evitar que esos sentimientos crezcan y evitar las recaídas.
Recaída mental
En la recaída mental hay una guerra en tu mente. Parte de ti quiere usar, pero parte de ti no. En la fase temprana de la recaída mental, simplemente estás pensando en usar. Pero en la fase posterior definitivamente estás pensando en usar.
Los signos de recaída mental son:
Pensar en las personas, los lugares y las cosas con las que consumiste
Idealizando tu consumo pasado
Mentir
Salir con viejos amigos de consumo
Fantaseando sobre el consumo
Pensando en recaer
Planificación de su recaída en torno a los horarios de otras personas.
Se hace más difícil tomar las decisiones correctas a medida que el impulso de la adicción se hace más fuerte.
Técnicas para el tratamiento de las necesidades mentales.
Reproduce la cinta a través. Cuando piensas en tomar, la fantasía es que podrás controlar tu consumo esta vez. Sólo tendrás una bebida. Pero pon la cinta a través. Una bebida generalmente lleva a más bebidas. Te despertarás al día siguiente sintiéndote decepcionado de ti mismo. Es posible que no puedas detenerte al día siguiente y te verás atrapado en el mismo círculo vicioso. Cuando reproduce esa cinta hasta su conclusión lógica, el uso no parece tan atractivo.
Un impulso mental común es que puede salirse con la suya, porque nadie sabrá si tiene una recaída. Tal vez su cónyuge esté ausente el fin de semana, o usted esté ausente en un viaje. Ahí es cuando su adicción intentará convencerlo de que no tiene un gran problema y que realmente está haciendo su recuperación para complacer a su cónyuge o su trabajo. Reproduce la cinta a través. Recuérdese las consecuencias negativas que ya ha sufrido y las posibles consecuencias que están a la vuelta de la esquina si recae nuevamente. Si pudieras controlar tu uso, ya lo habrías hecho.
Dígale a alguien que está teniendo deseos de consumir. Llame a un amigo, a un representante o a alguien en recuperación. Comparte con ellos lo que estás pasando. La magia de compartir es que en el momento en que empiezas a hablar sobre lo que estás pensando y sintiendo, tus impulsos comienzan a desaparecer. No parecen tan grandes y no te sientes tan solo.
Distraerse. Cuando pienses en consumir, haz algo para ocuparte. Llamar a un amigo. Ir a una reunión. Levántate y ve a pasear. Si simplemente te sientas allí con tu impulso y no haces nada, estás dando a tu recaída mental un lugar para crecer.
Espere 30 minutos. La mayoría de los impulsos generalmente duran menos de 15 a 30 minutos. Cuando estás en un impulso, se siente como una eternidad. Pero si puedes mantenerte ocupado y hacer las cosas que se supone que debes hacer, rápidamente desaparecerá.
Haga su recuperación un día a la vez. No pienses si puedes permanecer abstinente para siempre. Ese es un pensamiento paralizante. Es abrumador incluso para las personas que han estado en recuperación durante mucho tiempo.
Un día a la vez, significa que debe hacer coincidir sus objetivos con su fuerza emocional. Cuando se sienta fuerte y esté motivado para no consumir, entonces dígase que no consumirá para la próxima semana o el próximo mes. Pero cuando esté luchando y tenga muchas ganas, y esos momentos sucederán con frecuencia, dígase que no consumirá para hoy ni para los próximos 30 minutos. Recupérese en trozos pequeños y no se sabotee pensando demasiado hacia adelante.
Haga que la relajación sea parte de su recuperación. La relajación es una parte importante de la prevención de recaídas, porque cuando estás tenso, tiendes a hacer lo que es familiar e incorrecto, en lugar de lo que es nuevo y correcto. Cuando estás tenso, tiendes a repetir los mismos errores que hiciste antes. Cuando estás relajado estás más abierto al cambio.
Recaída física
Una vez que empiezas a pensar en una recaída, si no utilizas algunas de las técnicas mencionadas anteriormente, no tardarás mucho en ir a una recaída física: ir a tomar algo; llamar a su vendedor, por ejemplo, para opioides, cocaína o marihuana.
Es difícil detener el proceso de recaída en ese punto. Ahí no es donde debes enfocar tus esfuerzos en la recuperación. Eso es lograr la abstinencia a través de la fuerza bruta. Pero no es recuperación. Si reconoce los signos de advertencia tempranos de recaída y comprende los síntomas de la abstinencia postaguda, podrá recuperarse antes de que sea demasiado tarde.
Referencias
Las etapas de la recaída fueron descritas por primera vez por Terence Gorski. Gorski, T., & Miller, M., Staying Sober: A Guide for Relapse Prevention: Independence Press, 1986.
National Institute on Drug Abuse, National Institutes of Health, United States
Resumen: En los drogadictos abstinentes, las señales que antes se asociaban con las experiencias de consumo de drogas ganan potencia inductora de recaída (‘incubar‘) a través del tiempo. Los modelos animales de incubación pueden ayudar a desarrollar tratamientos para prevenir la recaída, pero estos modelos se han centrado de manera ubicua en el papel de los estímulos condicionados (CSs) para la administración de fármacos. Los estímulos discriminatorios (DS) son únicos porque ejercen un control de estímulo tanto en el consumo de drogas como en el comportamiento de búsqueda de drogas y son difíciles de extinguir. Por esta razón, la incubación de los efectos excitadores de los DS que indican la disponibilidad de medicamentos, aún no examinados en estudios preclínicos, podría ser relevante para la prevención de recaídas. Entrenamos ratas para autoadministrarse cocaína (o comida sabrosa) bajo el control de DS, y luego investigamos la incubación controlada por DS de los antojos, en ausencia de CS emparejados con drogas. La cocaína controlada por DS (pero no los alimentos sabrosos) buscó incubarse durante 60 días de abstinencia y persistió hasta 300 días. Comprender los mecanismos neuronales de esta incubación controlada por DS es prometedor para los tratamientos de recaída de drogas.
Más del 85% de las personas que abandonan una droga adictiva comienzan a usarla nuevamente dentro de un año. Las tasas de recaída han cambiado poco en las últimas cinco décadas. Las situaciones, lugares y objetos asociados con el consumo de drogas pueden provocar una recaída mucho después de la última exposición de una persona a una droga.
Podemos estudiar la recaída entrenando animales para autoadministrarse drogas como la cocaína. Por ejemplo, las ratas pueden aprender a presionar una palanca para recibir una infusión de una droga emparejada con una señal (un estímulo condicionado), como un tono o una luz. Después del entrenamiento, las ratas continúan presionando la palanca para buscar la droga, incluso si este comportamiento ya no la libera. Además, la presión de la palanca en respuesta a las señales aumenta con el tiempo durante varios meses después de su última exposición a la droga. Este fenómeno, conocido como incubación del deseo por las drogas, refleja el aumento en los antojos reportados por los consumidores de drogas abstinentes.
En los consumidores de drogas, estímulos como la pipa de crack o la jeringa usada para consumir la droga pueden contribuir más tarde a la recaída de la droga durante la abstinencia. La mayoría de los estudios que modelan este fenómeno se han centrado en cómo las ratas responden a un estímulo condicionado que señaló la administración de un medicamento durante el entrenamiento. Sin embargo, un segundo tipo de señal, conocido como estímulo discriminatorio, también puede influir en la recaída. Los estímulos discriminatorios son conjuntos de señales que indican si las drogas están a punto de estar disponibles o no; por ejemplo, la presencia de personas que venden drogas en una esquina frente a la presencia de la policía.
Madangopal et al. muestran ahora que los estímulos discriminativos, en ausencia de estímulos condicionados, también pueden controlar la incubación del deseo de drogas. Las ratas aprendieron a presionar una palanca en respuesta a una luz que indicaba la disponibilidad de cocaína (el estímulo discriminativo positivo), y a evitar responder a una luz diferente que indicara que la cocaína no estaba disponible (el estímulo discriminativo negativo). Cuando se probaron durante la abstinencia, las ratas solo aumentaron su palanca presionando a la primera luz con el tiempo, es decir, mostraron una incubación de ansia de drogas controlada por el estímulo discriminativo positivo. La presión de la palanca alcanzó su punto máximo después de 60 días de abstinencia y persistió hasta 300 días (casi la mitad de la vida útil de las ratas). Por el contrario, los mismos estímulos discriminativos no activaron el aumento de la presión de la palanca cuando se utilizaron para indicar la disponibilidad de un alimento sabroso.
Los estímulos discriminatorios son, por lo tanto, poderosos y persistentes desencadenantes del deseo por las drogas adictivas. Señalan la disponibilidad de una droga antes de consumir y recaer, lo que los convierte en un objetivo crítico para las estrategias de intervención. La comprensión de los mecanismos por los cuales los estímulos discriminativos promueven el deseo por las drogas podría conducir a nuevos tratamientos para prevenir la recaída.
Introducción
El riesgo de recaída es un obstáculo importante para el tratamiento eficaz de la adicción a las drogas (O’Brien, 2005; Wikler, 1973). En los consumidores abstinentes de drogas, varios factores contribuyen a la recaída, incluida la exposición a estímulos y contextos previamente asociados con el consumo de drogas (O’Brien et al., 1992), factores estresantes (Sinha, 2001) o la exposición aguda a la droga en sí (Jaffe et al., 1989). Los estudios preclínicos han recapitulado estos efectos en modelos de recaída utilizando ratones, ratas y primates no humanos (Venniro et al., 2016; Weiss, 2010). Un hallazgo importante en estos estudios es que la búsqueda de drogas inducida por estímulos (en ausencia de la droga) aumenta progresivamente durante la abstinencia, un fenómeno denominado incubación de ansia de drogas (Grimm et al., 2001; Neisewander et al., 2000). Se han demostrado aumentos de la búsqueda de drogas dependientes del tiempo en cocaína (Lu et al., 2004a), heroína (Shalev et al., 2001), metanfetamina (Shepard et al., 2004), alcohol (Bienkowski et al., 2004), y la nicotina (Abdolahi et al., 2010), así como las recompensas no farmacológicas como la sacarosa (Grimm et al., 2002). Estos hallazgos en roedores reflejan la incubación del ansia de drogas inducida por estímulos y las respuestas fisiológicas en adictos humanos (Bedi et al., 2011; Li et al., 2015a; Wang et al., 2013; Parvaz et al., 2016), y han sido importantes para el estudio de los mecanismos neuronales que contribuyen a la recaída de drogas (Dong et al., 2017; Marchant et al., 2013; Pickens et al., 2011; Wolf, 2016).
Los modelos de incubación preclínica han demostrado cómo los estímulos presentados después de la respuesta al consumo de la droga y el suministro de drogas durante el entrenamiento potencian la búsqueda de drogas cuando se presentan de forma contingente durante la abstinencia. Estos estímulos condicionados «confirmatorios» (CS) informan al animal de laboratorio que la respuesta al consumo de drogas se completó durante el entrenamiento. Los primeros estudios preclínicos de la incubación mostraron que también podría ocurrir en ausencia de tales CSs pareados con drogas (Lu et al., 2004a; Grimm et al., 2002). Esto sugiere que la incubación también podría ser inducida por otros estímulos asociados con el consumo de drogas, como las señales contextuales (por ejemplo, la cámara utilizada para el entrenamiento operante) o los estímulos discriminativos (DS) que indican la disponibilidad de medicamentos. (por ejemplo, la cámara utilizada para el entrenamiento operante) o estímulos discriminativos (DS) que indican la disponibilidad de medicamentos (por ejemplo, la luz de la casa que se ilumina durante la sesión de entrenamiento, la palanca retráctil que sirve como manipulandum operante). Sorprendentemente, se sabe poco acerca de los factores que subyacen a la incubación en ausencia de CS previamente emparejados con drogas. Un estudio reciente sugirió que no está mediado por señales contextuales (Adhikary et al., 2017), dejando a los DS como un probable culpable. Los DS son diferentes de las señales típicamente investigadas en estos estudios en que no son contingentes a la respuesta como las CS, ni están siempre presentes como señales contextuales. Más bien, los DS señalan la disponibilidad de drogas, o la falta de disponibilidad, lo que precede y guía el desempeño del comportamiento de consumo de drogas. Estudios anteriores han demostrado que una señalización de la disponibilidad de drogas DS (DS +) puede promover un comportamiento persistente de búsqueda de drogas mientras que una señalización de falta de disponibilidad de drogas DS (DS-) puede inhibir el comportamiento de consumo de drogas y el restablecimiento de la búsqueda de drogas inducido por el cebado de drogas (Weiss, 2010; Ettenberg, 1990; Gutman et al, 2017; Katner et al., 1999; McFarland y Ettenberg, 1997 ; Mihindou et al, 2013.; Yun and Fields, 2003; Pitchers et al., 2017). Además, el control de la búsqueda de drogas por DS persiste durante muchos meses y es altamente resistente a la extinción (Ciccocioppo et al., 2004; Ghitza et al., 2003; Martin-Fardon y Weiss, 2017). A pesar de la importancia de los DS en el control de estímulos de la toma y recaída de drogas, se desconoce si la búsqueda de drogas controlada por DS se incuba durante la abstinencia.
En este estudio, tratamos de evaluar directamente la contribución de los DS a la incubación, en ausencia de CS asociados con drogas. Con este fin, primero diseñamos un procedimiento basado en ensayos para entrenar ratas macho y hembras para autoadministrarse discriminativamente la cocaína (0,75 mg / kg / infusión) durante los ensayos en los que un DS + señaló la disponibilidad de cocaína, y para suprimir la respuesta en la misma palanca durante los ensayos en los que un DS señaló la falta de disponibilidad de cocaína durante la misma sesión. Las infusiones de drogas no se asociaron con CSs. Luego probamos la capacidad de los DS para controlar la búsqueda de cocaína en múltiples puntos de tiempo que se extienden hasta 400 días de abstinencia. Además, después de un cese completo del comportamiento de búsqueda de cocaína, evaluamos si una dosis de cebado restablecería la búsqueda de cocaína controlada por DS en las mismas ratas. Finalmente, para determinar si la incubación controlada por DS en nuestras condiciones experimentales era específica para la cocaína, entrenamos a un grupo separado de ratas en un procedimiento análogo que utilizaba alimentos sabrosos (45 mg de bolitas de alto contenido de carbohidratos) como recompensa operante y evaluamos el curso temporal de Búsqueda de alimentos controlada por DS.
Resultados
Experimento 1: incubación de búsqueda selectiva de cocaína controlada por estímulos
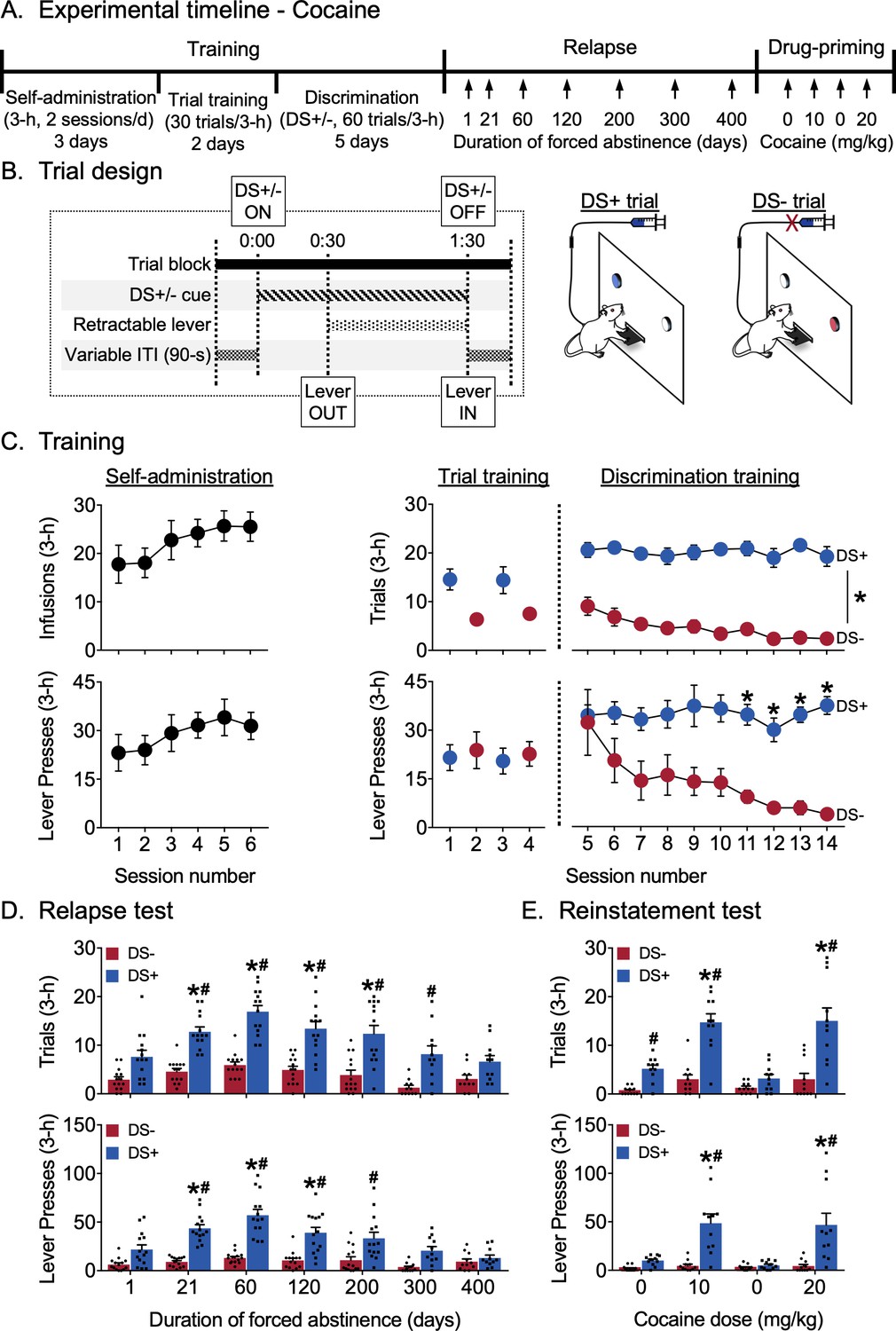
El objetivo del Experimento 1 era determinar la persistencia de la búsqueda de cocaína discriminada no reforzada (recaída en la búsqueda de cocaína controlada por DS) y evaluar la potenciación de esta respuesta de búsqueda durante la abstinencia (incubación de búsqueda de cocaína controlada por DS). Entrenamos a ratas macho y hembra para presionar una palanca retráctil central solo durante las pruebas en las que la entrada de la palanca fue precedida por la iluminación de un estímulo de luz que señalaba la disponibilidad de cocaína (pruebas DS +) y para suprimir la respuesta durante las pruebas cuando la disponibilidad de la misma palanca fue precedida por un segundo estímulo de luz que indica ausencia de recompensa de cocaína (ensayos DS). No hubo señales discretas adicionales emparejadas con recompensas. Una vez capacitados, utilizamos un diseño dentro de los sujetos para evaluar la búsqueda discriminada de cocaína (condiciones de extinción) después de diferentes duraciones de abstinencia que se extienden hasta 400 días. Después del cese completo del comportamiento de búsqueda de cocaína en el día 400 de abstinencia, Utilizamos un diseño dentro de los sujetos y un procedimiento de dosis-respuesta de cocaína ascendente para evaluar la capacidad de preparar inyecciones de cocaína para restablecer la búsqueda de cocaína controlada por DS. Todos los datos de comportamiento pertenecientes al Experimento 1 se muestran enFigura 1 (colapsada por sexo) y Figura 1: suplemento de figura 1 (desglosado por sexo). Los resultados estadísticos para todos los análisis relacionados con el experimento se proporcionan en formato tabular como en la Figura 1: fuente de datos 1.
Figura 1 con 2 suplementos
Incubación de la búsqueda discriminativa de cocaína controlada por estímulos. (A) Cronología experimental. (B) Esquema que muestra el momento de los eventos individuales durante un solo ensayo de DS de 3 minutos, y las diferencias entre los dos tipos de ensayos durante el entrenamiento de discriminación para la recompensa de cocaína. Las ratas recibieron recompensa de cocaína (0,75 mg / kg / infusión) durante los ensayos DS + pero no recibieron recompensa de cocaína durante los ensayos DS (n = 16). (C) Datos de entrenamiento. Autoadministración: las ratas aprendieron a autoadministrarse cocaína durante seis sesiones. Número medio (± SEM) de infusiones de cocaína y prensas de palanca durante cada sesión de 3 horas. Entrenamiento de prueba: número medio (± SEM) de pruebas DS + o DS- con al menos una palanca de presión (denotado como pruebas), y el número de prensas de palanca durante las sesiones de 3 h (denotadas como prensas de palanca) con 30 ensayos de un tipo de ensayo único (ensayos DS + en la sesión AM, ensayos DS en la sesión PM). Entrenamiento de discriminación: en más de 10 sesiones, las ratas aprendieron a discriminar DS + de los ensayos de DS. Número medio (± SEM) de ensayos y presiones de palanca durante la sesión de entrenamiento de discriminación de 3 horas con 30 ensayos cada uno de los ensayos DS + y DS- presentados de forma pseudoaleatoria. * indica una diferencia significativa (p <0.05) entre responder durante los tipos DS + y DS-trial (n = 14). (D) Prueba de recaída. La incubación de la palanca que responde durante DS +, pero no DS-, los ensayos alcanzaron su punto máximo a los 60 días de abstinencia y regresaron a los niveles basales durante 400 días. Número medio (± SEM) de ensayos y presiones de palanca durante las sesiones de prueba de recaída de 3 horas (30 ensayos de DS + y DS- presentados de forma pseudoaleatoria) en condiciones de extinción. * denota una diferencia significativa (p <0.05) de responder durante el día 1. Las columnas indican la media (± SEM) para el grupo, mientras que los puntos indican valores para ratas individuales. # denota una diferencia significativa (p <0.05) entre DS + y DS-respondiendo durante las sesiones de prueba (n = 11–14). (E) Prueba de restablecimiento. Las ratas restablecieron la búsqueda de cocaína controlada por DS en respuesta a inyecciones IP de cocaína (10 y 20 mg / kg), pero no solución salina. Número medio (± SEM) de ensayos y prensas de palanca durante las 3 horas de sesiones de prueba de restablecimiento preparadas con solución salina o cocaína (30 ensayos de DS + y DS- presentados de manera pseudoaleatoria) en condiciones de extinción. Las columnas indican la media (± SEM) para el grupo, mientras que los puntos indican valores para ratas individuales. * denota una diferencia significativa (p <0.05) de responder en la primera sesión de prueba de solución salina (dosis de cocaína = 0 mg / kg). # denota una diferencia significativa (p <0.05) entre DS + y DS-respondiendo durante la sesión de prueba (n = 11). Ver Figura 1: suplemento de figura 1para datos de comportamiento del experimento, desglosados por sexo. Ver Figura 1: suplemento de figura 2 para los pesos corporales del sujeto, desglosados por sexo. Consulte la Figura 1: fuente de datos 1 para ver una tabla de resultados estadísticos relacionados con el experimento. Figura 1: fuente de datos 1 Producto estadístico para el Experimento 1: Incubación de la búsqueda discriminativa de cocaína controlada por estímulos (los análisis correspondientes a la Figura 1 se resaltan en gris).
Formación
La línea de tiempo experimental y diseño de los ensayos individuales se muestran en la Figura 1A, B. Las ratas aprendieron a responder en la palanca de recompensa de cocaína durante las primeras seis sesiones de acceso continuo (Figura 1C). Continuaron respondiendo en el formato de prueba y luego aprendieron a discriminar DS + de DS- durante el entrenamiento de discriminación. Se analizó el número de pruebas ‘exitosas’ (denotadas como pruebas y definidas como hacer al menos una presión de palanca durante una prueba) y la cantidad total de prensas de palanca (denotadas como prensas de palanca y registradas por separado para cada tipo de prueba DS) durante cada sesión. Utilizamos un modelo multinivel basado en la probabilidad máxima de dos vías con factores dentro de los sujetos Sesión (sesiones de discriminación 5–14) y DS (DS +, DS-). En los ensayos, observamos un efecto principal significativo de la DS (F 1,13 = 948.21, p <0.0001) pero no de la Sesión, y no hubo interacción entre la Sesión y la DS, lo que indica que la respuesta durante las pruebas DS + fue mayor que la respuesta durante las pruebas DS durante todas las sesiones de entrenamiento en discriminación. Para las prensas de palanca, observamos un efecto principal significativo de DS (F 1,13 = 161.63, p <0.0001) y una interacción entre Session y DS (F 9,117 = 2.62, p = 0.0085) pero ningún efecto principal de Session. Los análisis post-hoc indicaron que la respuesta durante los ensayos de DS + fue mayor que la respuesta en los ensayos de DS durante las últimas cuatro sesiones de entrenamiento de discriminación (p <0.05).
Prueba de recaída
La Figura 1D muestra una recaída en términos de respuesta media durante 3 horas de sesiones de prueba de discriminación no reforzada para la búsqueda de cocaína. Se probaron las mismas ratas en diferentes puntos de tiempo 1-400 días después del entrenamiento de discriminación. Al igual que durante el entrenamiento, analizamos las pruebas y las medidas de prensas de palanca. Utilizamos un modelo factorial bidireccional con factores dentro de los sujetos de días de abstinencia forzada (1, 21, 60, 120, 200, 300 y 400 días) y tipo de DS (DS +, DS-). Para los ensayos , observamos efectos principales significativos de Día (F 6,72 = 9.68, p <0.0001) y DS (F 1,13 = 257.53, p <0.0001) y una interacción entre los dos (F 6,72= 4.30, p = 0.0009), lo que refleja un mayor número de ensayos ‘exitosos’ asociados con la presentación de DS + después de 21, 60, 120 y 200 días de abstinencia en comparación con el día de abstinencia (p <0.05) y más ensayos ‘exitosos’ de DS + comparados a los ensayos de DS en los días de abstinencia 21, 60, 120, 200 y 300 (p <0,05). El número de ensayos de DS «exitosos» no aumentó significativamente a lo largo de los días. Para las prensas de palanca , observamos efectos principales significativos de Día (F 6,72 = 8.94, p <0.0001) y DS (F 1,13 = 182.25, p <0.0001), y una interacción entre los dos factores (F 6,72= 7.95, p <0.0001), lo que refleja un mayor número de prensas de palanca asociadas con la presentación de DS + después de 21, 60 y 120 días de abstinencia en comparación con el día de abstinencia (p <0.05) y una mayor respuesta durante los ensayos de DS + en comparación con los ensayos de DS en los días de abstinencia 21, 60, 120 y 200 (p <0.05). El número de prensas de palanca durante los ensayos DS no aumentó significativamente durante los días. En general, los ensayos datos indican incubación de ensayos ‘exitosos’ durante DS +, pero no DS-, ensayos después de 21-200 días de abstinencia, mientras que las prensas de palanca datos indican incubación del número de prensas de palanca durante DS +, pero no DS-, ensayos después de 21-120 días de abstinencia. Además, las ratas mantuvieron una respuesta discriminatoria hasta 300 días (según la medida de prueba) o 200 días (según la medida de prensas de palanca) después de la última sesión de entrenamiento.
Prueba de restablecimiento
La Figura 1E muestra el restablecimiento en términos de respuesta media durante una sesión de discriminación no reforzada de 3 horas para la búsqueda de cocaína después de cebar inyecciones de cocaína o solución salina. Para ambos ensayos y medidas de prensas de palanca, utilizamos un modelo factorial bidireccional con los factores dentro de los sujetos. Condición de tratamiento (solución salina 1, 10 mg / kg de cocaína, solución salina 2, 20 mg / kg de cocaína) y tipo DS (DS +, DS -). Para los ensayos , hubo efectos principales significativos del tratamiento (F 3,30 = 15.35, p <0.0001) y DS (F 1,10 = 108.66, p <0.0001) y una interacción entre los dos factores (F 3,30= 12.42, p <0.0001), lo que refleja un mayor número de ensayos «exitosos» asociados con la presentación de DS + después de ambas dosis de cebado de cocaína (10 y 20 mg / kg) en comparación con la solución salina y una mayor respuesta durante los ensayos de DS + en comparación con los ensayos de DS después de ambas cocaína dosis de cebado, así como después de la primera inyección de cebado salino (p <0.05). El número de ensayos «exitosos» asociados con la presentación de DS no fue alterado por las condiciones del tratamiento. Para las prensas de palanca , observamos efectos principales significativos del tratamiento (F 3,30 = 8.31, p = 0.0004) y DS (F 1,10 = 45.73, p <0.0001) y una interacción entre los dos factores (F 3,30= 9,45, p = 0,0001), lo que refleja un mayor número de prensas de palanca durante los ensayos DS + después de las dos inyecciones de cebado de cocaína (10 y 20 mg / kg) en comparación con la solución salina (Saline1 y Saline2), así como una mayor respuesta durante los ensayos DS + en comparación con Pruebas de DS durante las dos inyecciones de cebado de cocaína (p <0.05). El número de prensas de palanca asociadas con la presentación DS no fue alterado por las condiciones del tratamiento. En general, los datos de ensayos y prensas de palanca indicaron un restablecimiento confiable cebado con cocaína durante DS +, pero no DS-, ensayos que ocurrieron más de 400 días después de la última sesión de entrenamiento de discriminación.
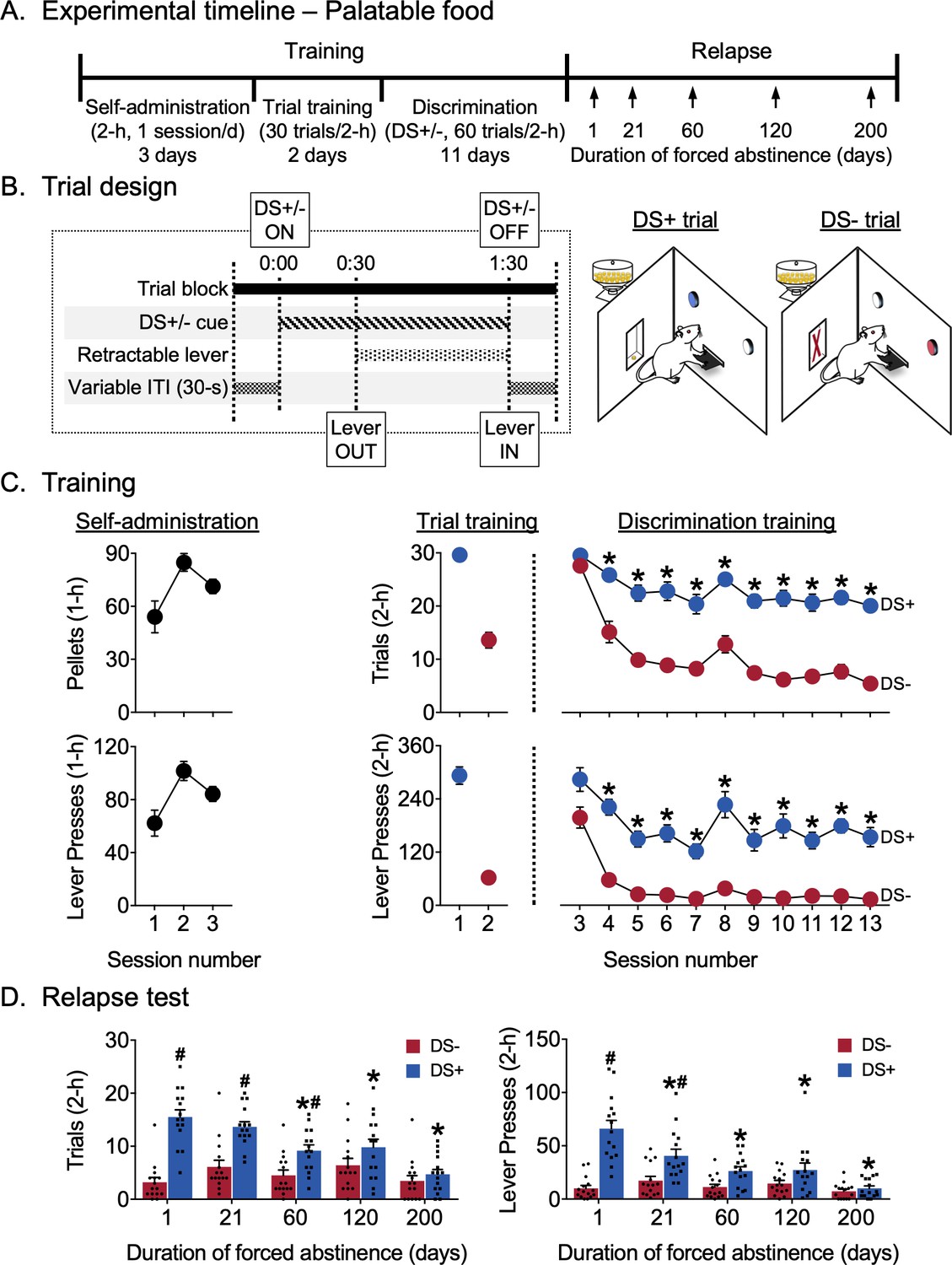
Experimento 2: reducción de la búsqueda de alimentos sabrosos controlados por estímulos discriminativos
El objetivo del Experimento 2 era determinar si la persistencia y potenciación de la búsqueda controlada por DS observada en el Experimento uno se generalizaría a una recompensa de comida aceptable. Entrenamos a ratas machos y hembras para presionar con palanca para obtener una recompensa de alimentos sabrosos usando un procedimiento de entrenamiento similar al del Experimento 1. Después del entrenamiento, utilizamos un diseño dentro de los sujetos para probar las ratas en busca de alimentos sabrosos discriminados después de diferentes duraciones de extensión Hasta 200 días. Todos los datos de comportamiento pertenecientes al Experimento 2 se muestran en la Figura 2 (colapsada en función del sexo) y en la Figura 2: suplemento de figura 1 (desglosado por sexo). Los resultados estadísticos para todos los análisis relacionados con el experimento se proporcionan en formato tabular como en la Figura 2: fuente de datos 1 .
Figura 2 con 3 suplementos
Disminución de la búsqueda discriminativa de alimentos sabrosos controlados por estímulos. (A) Cronología experimental. (B) Esquema que muestra el momento de los eventos individuales durante un único ensayo de DS de 2 minutos, y las diferencias entre los dos tipos de ensayos durante el entrenamiento de discriminación para la recompensa de alimentos apetecibles (45 mg de pastillas de carbohidratos altos). Las ratas recibieron recompensa alimentaria durante los ensayos DS + pero no recibieron recompensa durante los ensayos DS (n = 16). (C) Datos de entrenamiento. Autoadministración: las ratas aprendieron a autoadministrarse alimentos sabrosos durante tres sesiones. Número medio (± SEM) de gránulos de comida sabrosa recibidos y prensas de palanca durante cada sesión de 1 hora. Entrenamiento de prueba: número medio (± SEM) de pruebas DS + o DS- con al menos una palanca de presión (denotado como pruebas), y el número de prensas de palanca durante las sesiones de 2 h (denotadas como prensas de palanca) con 30 ensayos de un solo tipo de ensayo (ensayos DS + en la sesión AM, ensayos DS en la sesión PM). Entrenamiento de discriminación: durante 11 sesiones, las ratas aprendieron a discriminar DS + de los ensayos de DS. Número medio (± SEM) de ensayos y prensas de palanca durante la sesión de entrenamiento de discriminación de 2 horas con 30 ensayos cada uno de los ensayos DS + y DS- presentados de manera pseudoaleatoria. * indica una diferencia significativa (p <0.05) entre la respuesta durante los ensayos DS + y DS- (n = 15). (D) Prueba de recaída. Palanca respondiendo durante DS +, pero no DS-, los ensayos alcanzaron su punto máximo a 1 día de abstinencia y disminuyeron durante 200 días. Número medio (± SEM) de ensayos y prensas de palanca durante las sesiones de prueba de recaída de 2 horas (30 ensayos de DS + y DS- presentados de forma pseudoaleatoria) en condiciones de extinción. * denota una diferencia significativa (p <0.05) de responder durante el día 1. Las columnas indican la media (± SEM) para el grupo, mientras que los puntos indican valores para ratas individuales. # denota una diferencia significativa (p <0.05) entre DS + y DS-respondiendo durante la prueba (n = 15). Consulte la Figura 2: figura 1 del suplemento para ver los datos de comportamiento del experimento, desglosados por sexo. Consulte la Figura 2: figura 2 del suplemento para obtener las recompensas de alimentos obtenidas durante el entrenamiento de discriminación, desglosadas por sexo. Consulte la Figura 2: suplemento de la figura 3 para conocer los pesos corporales de los sujetos, desglosados por sexo. VerFigura 2: fuente de datos 1 para una tabla de resultados estadísticos relacionados con el experimento. Figura 2: datos de origen 1 Producto estadístico para el Experimento 2: Reducción de la búsqueda de alimentos sabrosos controlados por estímulos discriminativos (los análisis relacionados con la Figura 2 se resaltan en gris).
Formación
Las ratas aprendieron a responder en la palanca de la recompensa de comida sabrosa durante las primeras tres sesiones de acceso continuo ( Figura 2C ). Las ratas continuaron respondiendo en el formato de prueba y luego aprendieron a discriminar DS + de DS- durante el entrenamiento de discriminación. Para el análisis de la discriminación exitosa en ambos ensayos y medidas de prensas de palanca, utilizamos un modelo factorial bidireccional con factores dentro de los sujetos Sesión (sesiones de entrenamiento de discriminación 3-13) y DS (DS +, DS-). Para los ensayos, observamos efectos principales significativos de Sesión (F 10,140 = 15.42, p <0.0001) y DS (F 1,14 = 1014.94, p <0.0001) y una interacción entre los dos factores (F 10,140= 8,22, p <0,0001), lo que refleja una mayor respuesta durante los ensayos DS + para todos excepto la primera sesión (p <0,05). Para las prensas de palanca, observamos efectos principales significativos de Session (F 10,140 = 15.63, p <0.0001) y DS (F 1,14 = 577.71, p <0.0001) y una interacción entre los dos factores (F 10,140 = 2.31, p = 0.0151), que nuevamente refleja una mayor respuesta durante los ensayos DS + para todos menos la primera sesión (p <0.05).
Prueba de recaída
La Figura 2D muestra la recaída en términos de respuesta media durante las sesiones de prueba de discriminación no reforzada de 2 horas para la búsqueda de alimentos sabrosos. Se probaron las mismas ratas en diferentes puntos de tiempo 1–200 días después del entrenamiento de discriminación. Al igual que durante el entrenamiento, analizamos las pruebas y las medidas de prensas de palanca. Para ambos ensayos y medidas de prensas de palanca, utilizamos un modelo factorial bidireccional con factores dentro de los sujetos de días de abstinencia forzada (1, 21, 60, 120 y 200 días) y tipo de DS (DS +, DS-). Para los ensayos , observamos efectos principales significativos de Day (F 4,56 = 5.57, p = 0.0008) y DS (F 1,14 = 133.04, p <0.0001) y una interacción entre los dos factores (F4,56 = 14.29, p <0.0001), lo que refleja un menor número de ensayos ‘exitosos’ asociados con la presentación de DS + después de 60, 120 y 200 días de abstinencia que después de un día de abstinencia (p <0.05) y mayor respuesta a DS + que DS-on abstinencia días 1, 21 y 60 (p <0,05). El número de ensayos «exitosos» asociados con la presentación de DS no aumentó con los días. Para las prensas de palanca, observamos efectos principales significativos del Día de la abstinencia (F 4,56 = 8,57, p <0,0001) y DS (F 1,14 = 101,92, p <0,0001) y una interacción entre los dos (F 4,56= 17.44, p <0.0001), que refleja un menor número de prensas de palanca asociadas con la presentación de DS + después de 21, 60, 120 y 200 días de abstinencia que después de un día de abstinencia (p <0.05) y mayor respuesta a DS + que DS- en los días de abstinencia 1 y 21 (p <0,05). El número de prensas de palanca asociadas con la presentación DS no cambió con los días. En general, los datos de ensayos y prensas de palanca indican que la búsqueda de alimentos disminuyó o disminuyó con el tiempo y que las ratas mantuvieron una respuesta discriminativa durante solo 60 días (según la medida de los ensayos) o 21 días (según la medida de prensas de palanca) después de la última sesión de entrenamiento.
Discusión
Utilizamos un procedimiento de prueba para investigar la incubación de la cocaína o la búsqueda de alimentos sabrosos controlados por estímulos discriminativos que señalen la disponibilidad (DS +) o la indisponibilidad (DS-) de las recompensas en ausencia de CS de recompensa asociada. Las ratas aprendieron fácilmente a responder al DS + para la cocaína (Experimento 1) o para el alimento (Experimento 2) e inhibir la respuesta al DS en la misma sesión. La búsqueda de cocaína controlada por DS fue máxima después de 60 días de abstinencia (lo que refleja la incubación de la búsqueda de cocaína controlada por DS) y persistió hasta 300 días. Además, cuando la búsqueda de cocaína controlada por DS se extinguió por completo después de 400 días de abstinencia, las inyecciones de cebado de cocaína restablecieron la búsqueda de cocaína. En contraste, la búsqueda de alimentos controlada por DS fue máxima a 1 día de abstinencia, disminuyó progresivamente con el tiempo y ya no se observó después de 60 días de abstinencia. Por lo tanto, la incubación de la búsqueda de recompensas controlada por DS en nuestras condiciones experimentales fue específica para la cocaína.
En estudios anteriores, se ha demostrado que los DS combinados con la autoadministración de cocaína promueven la búsqueda de drogas que es altamente resistente a la extinción en múltiples sesiones de pruebas no reforzadas (Ciccocioppo et al., 2004; Martin-Fardon y Weiss, 2017; Weiss et al., 2000). Las entregas de recompensa en estos estudios se combinaron con CS discretas adicionales, y los DS contrastantes se combinaron con diferentes palancas y se presentaron en sesiones separadas, lo que dificulta la separación de la contribución potencial de los DS de los CS y los estímulos contextuales. En nuestros experimentos, las entregas de recompensa no se emparejaron con CS adicionales durante el entrenamiento, y los dos DS contrastantes se emparejaron con una palanca retráctil común y se presentaron en un orden pseudoaleatorizado dentro de la misma sesión. Después del entrenamiento, las ratas se probaron en condiciones no reforzadas para la búsqueda de drogas controlada por DS, utilizando el mismo programa de presentación de DS que durante el entrenamiento. Debido a que se requirió el mismo manipulandum operante y respuesta para buscar refuerzo en respuesta a cada DS dentro de la misma sesión de prueba, Sabemos que la búsqueda de drogas discriminada en nuestro modelo fue controlada exclusivamente por los DS y no por estímulos contextuales, señales espaciales clásicamente condicionadas, presentación del manipulandum operante o incluso el desempeño de la respuesta de búsqueda de drogas. Bajo estas condiciones, observamos una búsqueda persistente de medicamentos no reforzados durante las presentaciones de DS +, pero no las presentaciones de DS, hasta 300 días después del último emparejamiento de medicamentos de DS. Estos datos amplían estudios previos sobre la búsqueda de drogas controladas por DS y sugieren que además de establecer la ocasión para el comportamiento de búsqueda de drogas, el DS + es suficiente para motivar la búsqueda de drogas en ausencia de CS explícitos de drogas. o incluso el rendimiento de la respuesta de búsqueda de drogas. Bajo estas condiciones, observamos una búsqueda persistente de medicamentos no reforzados durante las presentaciones de DS +, pero no las presentaciones de DS, hasta 300 días después del último emparejamiento de medicamentos de DS. Estos datos amplían estudios previos sobre la búsqueda de drogas controladas por DS y sugieren que además de establecer la ocasión para el comportamiento de búsqueda de drogas, el DS + es suficiente para motivar la búsqueda de drogas en ausencia de CS explícitos de drogas. o incluso el rendimiento de la respuesta de búsqueda de drogas. Bajo estas condiciones, observamos una búsqueda persistente de medicamentos no reforzados durante las presentaciones de DS +, pero no las presentaciones de DS, hasta 300 días después del último emparejamiento de medicamentos de DS. Estos datos amplían estudios previos sobre la búsqueda de drogas controladas por DS y sugieren que además de establecer la ocasión para el comportamiento de búsqueda de drogas, el DS + es suficiente para motivar la búsqueda de drogas en ausencia de CS explícitos de drogas.
Durante mucho tiempo se ha apreciado en la investigación básica del comportamiento que el aprendizaje sobre las DS operantes implica tanto el condicionamiento clásico como el operante ( Mowrer, 1960 ; Rescorla y Solomon, 1967 ; Weiss, 1978 ; Weiss, 2014 ). Según lo explicado por Rescorla y Salomón (1967) Todas las condiciones necesarias para el condicionamiento clásico están presentes durante la respuesta operante discriminada. Por lo tanto, además de establecer la ocasión para la obtención de recompensas, debe esperarse que un DS + venga a provocar respuestas condicionadas clásicamente (CR). Tales RC pueden incluir la inducción de estados motivacionales (es decir, ansia de drogas). En el presente estudio, las ratas aprendieron a presionar con palanca durante el DS + para la cocaína (es decir, aprendieron una relación estímulo-respuesta-resultado y su comportamiento quedó bajo control del estímulo). Debido a que solo recibieron cocaína durante el DS +, también deberían haber aprendido una relación de estímulo-resultado pavloviano que imbuiría al DS + con propiedades motivacionales de incentivo ( Weiss, 2014). Estas propiedades motivacionales de excitación, adquiridas a través de procesos pavlovianos, probablemente contribuyeron al efecto de incubación observado en este estudio.
Demostramos que la búsqueda de cocaína controlada por DS se potencia durante la abstinencia (es decir, mostramos la incubación del deseo de cocaína controlado por DS) incluso en ausencia de CS explícitos de drogas. Los estudios de incubación han empleado típicamente procedimientos de prueba entre sujetos en los que ratas previamente entrenadas para autoadministrarse una droga adictiva se devuelven a las mismas cámaras después de diversos períodos de abstinencia y se analiza la búsqueda de drogas con o sin los CS previamente emparejados con la droga ( Pickens et al., 2011 ; Li et al., 2015b ; Lu et al., 2004b ). Después de la autoadministración de la cocaína, la respuesta a los CS emparejados con cocaína aumentó progresivamente («incubó») durante los primeros 60 a 90 días de retiro ( Grimm et al., 2001 ;Grimm et al., 2003 ). Sin embargo, la incubación también se ha observado en ausencia de CS emparejados con el fármaco; la extinción que responde a la ausencia de los CS emparejados con el medicamento también aumentó progresivamente durante hasta 90 días ( Grimm et al., 2003 ) y persistió hasta 180 días ( Lu et al., 2004a ). Un estudio reciente demostró que esta potenciación no está mediada por señales contextuales ( Adhikary et al., 2017). Sin embargo, los factores que controlan la incubación en ausencia de CS previamente emparejados con el fármaco no fueron aclarados. En el presente estudio, encontramos un aumento dependiente del tiempo en la búsqueda de medicamentos (incubación) durante las presentaciones de DS + en ausencia de cualquier CS explícito de medicamentos emparejados durante la abstinencia. La búsqueda controlada por DS continuó aumentando hasta los 60 días de abstinencia y persistió hasta 300 días (aproximadamente la mitad de la vida útil de una rata). Este curso temporal de búsqueda de cocaína controlada por DS es especialmente notable si se considera que el mismo grupo de ratas estuvo expuesto a pruebas repetidas de recaída en condiciones de extinción. Es posible que en un diseño entre sujetos, la incubación controlada por DS muestre una fase de aumento más prolongada que la observada aquí y persista más de 300 días, en ausencia de aprendizaje de extinción en pruebas de recaída repetidas.
En contraste, la búsqueda de cocaína en los ensayos de DS no se incubó; las ratas continuaron reprimiendo la respuesta en los ensayos de DS durante todas las pruebas de recaída y mantuvieron la discriminación hasta 300 días de abstinencia. Se ha demostrado que los DS que indican la falta de disponibilidad de la cocaína inhiben la autoadministración continua de la cocaína y suprimen el restablecimiento inducido por el cebado de la cocaína ( Mihindou et al., 2013). Desde la perspectiva de la traducción y el desarrollo del tratamiento, la inhibición de la búsqueda de cocaína puede ser tan importante como su potenciación. El comportamiento guiado por cada DS en nuestro estudio fue capaz de sobrevivir a múltiples sesiones de extinción y pruebas subsiguientes de reincorporación inducida por el cebado de cocaína – después de que la respuesta del DS + se extinguió a los niveles del DS – inyecciones de cebado de cocaína reinstaladas específicamente durante el DS +, pero -, ensayos. Los estudios futuros con este procedimiento disociarán los mecanismos neurobiológicos que permiten a estos dos DS funcionalmente ortogonales mediar la incubación de la búsqueda de cocaína controlada por DS.
Usando un formato similar de DS y presentación de palanca, también entrenamos ratas para presionar con palanca para obtener una comida sabrosa durante las pruebas de DS +, pero no de DS. Encontramos que las ratas con DS alimentario obtuvieron más respuestas totales que las ratas con cocaína DS durante el entrenamiento, mantuvieron su discriminación respondiendo en condiciones no reforzadas y también mostraron respuestas de búsqueda más altas que las ratas con cocaína-DS durante la prueba de recaída inicial en el día 1. Sin embargo , bajo el mismo programa de pruebas repetidas utilizado para la recaída de cocaína, extinguieron rápidamente su respuesta controlada por DS en ausencia de alimentos y disminuyeron progresivamente la búsqueda de alimentos durante la abstinencia. Es posible que la búsqueda de alimentos controlada por DS se hubiera incubado en ausencia de pruebas repetidas de recaída en extinción. De hecho, se ha observado la incubación utilizando el procedimiento clásico con recompensa de sacarosa oral;Lu et al., 2004b ; Grimm et al., 2003 ). Es poco probable que las diferencias en la búsqueda no farmacológica en respuesta a los CS y DS sean el resultado de la elección del reforzador no farmacológico, como un estudio reciente demostró la incubación de la búsqueda de recompensa inducida por CS utilizando los mismos gránulos de alimentos sabrosos ( Krasnova et al. ., 2014 ). La mayor persistencia de la búsqueda en respuesta a los DS de drogas sobre alimentos observada aquí es más probablemente el resultado de una diferencia inherente en la fuerza del estímulo-control ejercido por las drogas sobre alimentos-DS. Nuestros hallazgos están de acuerdo con estudios anteriores que comparan directamente los DS pareados de fármacos y alimentos ( Ciccocioppo et al., 2004 ; Martin-Fardon y Weiss, 2017) pero también de manera más general, con estudios que comparan CS combinados con medicamentos y alimentos ( Tunstall y Kearns, 2016 ; N. Kearns et al., 2011 ). Se requieren estudios futuros para determinar si esta divergencia de los efectos de la DS en la búsqueda de drogas versus alimentos se debe a las diferencias en la fuerza de las asociaciones iniciales de recompensa de la DS durante el entrenamiento o debido a neuroadaptaciones específicas de la droga que surgen durante la abstinencia ( Wolf, 2016 ; Grimm et al., 2003 ; Shaham and Hope, 2005 ).
En conjunto, los resultados de los experimentos actuales muestran que la búsqueda de drogas operantes controladas por DS se incuba durante la abstinencia prolongada y persiste hasta 300 días de abstinencia a pesar de las repetidas pruebas de recaída. Sin embargo, al utilizar un procedimiento de prueba repetido similar, observamos una reducción de la búsqueda de alimentos sabrosos controlada por DS. Como señalamos anteriormente, los comportamientos controlados por DS ofrecen un camino especialmente prometedor para el desarrollo del tratamiento porque los DS siempre están presentes antes y durante la toma de drogas por parte del ser humano; No se limitan a acompañarlo o seguirlo. Pueden jugar un papel crítico en la recaída; por ejemplo, un estudio que midió el deseo de fumar de las azafatas de vuelo mostró que el deseo se acercaba al final de los vuelos cuando se acercaba la oportunidad de fumar un cigarrillo, independientemente de la duración del vuelo o del tiempo transcurrido desde el último cigarrillo (Dar et al., 2010 ). Los modelos animales de otros aspectos del comportamiento adictivo han sido cuestionados, por nosotros y por otros autores, porque el momento o la secuencia de los eventos no refleja las experiencias típicas de los usuarios de drogas humanos ( Epstein y Kowalczyk, 2018 ; Vanderschuren et al., 2017 ). El procedimiento que describimos aquí aborda estas preocupaciones en el ámbito de la reactividad cue y su incubación, y es muy adecuado para separar el complejo conjunto de mecanismos de comportamiento y neurales que subyacen a las contribuciones de DSs a la recaída ( Bradfield y Balleine, 2013 ; Colwill y Rescorla, 1990 ; Rescorla, 1990 ; de Wit y Dickinson, 2009 ).
Materiales y métodos
Diseño experimental
El objetivo de este estudio fue evaluar la capacidad de los estímulos discriminativos que indican la disponibilidad de cocaína para potenciar la búsqueda de cocaína después de la abstinencia y luego determinar si este efecto se generalizaría a recompensas sin drogas. En la siguiente subsección se incluye una descripción detallada de los sujetos, aparatos y procedimientos experimentales. Primero proporcionamos una descripción general de los experimentos de comportamiento específicos.
Experimento 1: incubación de la búsqueda discriminativa de cocaína controlada por estímulos
El objetivo del Experimento 1 era determinar la persistencia de la búsqueda de cocaína discriminada no reforzada (recaída en la búsqueda de cocaína controlada por DS) y probar la potenciación de esta respuesta de búsqueda durante la abstinencia (incubación de la búsqueda de cocaína controlada por DS). La línea de tiempo del experimento se muestra en la Figura 1A. Entrenamos ratas machos y hembras utilizando dos sesiones diarias de 3 horas (mañana y tarde) para presionar una palanca retráctil central solo durante los ensayos en los que la entrada de la palanca fue precedida por la iluminación de un estímulo de luz que señalaba cocaína (0,75 mg / kg / infusión) disponibilidad (ensayos DS +) y para suprimir la respuesta durante los ensayos cuando la disponibilidad de la misma palanca fue precedida por un segundo estímulo de luz que indica ausencia de recompensa de cocaína (ensayos DS). No hubo señales discretas adicionales emparejadas con recompensas. Después de un entrenamiento exitoso (20 sesiones durante 10 días), probamos la búsqueda discriminada de cocaína usando un diseño dentro de los sujetos, después de que las duraciones de abstinencia variaran hasta los 400 días. Para pruebas de recaída, utilizamos el mismo procedimiento basado en pruebas y registramos el número de pruebas exitosas (definidas como hacer al menos una palanca de presión durante una prueba) y la cantidad total de prensas de palanca (registradas por separado para DS + y DS- pruebas) realizadas durante las 3 horas sesiones Finalmente, después del cese completo del comportamiento de búsqueda de cocaína en el día 400 de abstinencia, evaluamos la capacidad de preparar inyecciones de cocaína para restablecer la búsqueda de cocaína controlada por DS utilizando un diseño dentro de los sujetos y un procedimiento de dosis-respuesta de cocaína ascendente.
Experimento 2: disminución de la búsqueda de alimentos sabrosos, controlados por estímulos discriminativos
El objetivo del experimento dos era determinar si la persistencia y potenciación de la respuesta observada en el Experimento 1 se generalizaría a una recompensa no farmacológica. La línea de tiempo del experimento se muestra en la Figura 2A . Primero entrenamos ratas machos y hembras usando sesiones diarias de 2 horas (mañana o tarde) para presionar la palanca para obtener una recompensa de comida sabrosa (45 mg de pellets con alto contenido de carbohidratos) usando un procedimiento similar al utilizado en el Experimento 1. Después del entrenamiento exitoso (16 sesiones durante 16 días), probamos todas las ratas en busca de comida sabrosa discriminada usando un diseño dentro de los sujetos similar al del Experimento 1 pero usando sesiones de 2 horas, después de diferentes duraciones de abstinencia que se extienden hasta 200 días.
Asignaturas
Utilizamos ratas Sprague-Dawley macho (n = 16) y hembra (n = 16) (Charles River, EE. UU.; RRID: RGD_70508), con un peso de 250-350 g antes de la cirugía y el entrenamiento. En el experimento 1 con entrenamiento de autoadministración de cocaína, emparejamos ratas del mismo sexo durante 1 semana (n = 8 cada macho y hembra) antes de la cirugía y las alojamos individualmente después de la cirugía intravenosa, durante las fases de entrenamiento y abstinencia. En el Experimento 2 con entrenamiento de autoadministración de alimentos, emparejamos ratas del mismo sexo durante 1 semana (n = 8 cada macho y hembra) antes del comienzo del entrenamiento conductual y las alojamos individualmente durante el entrenamiento y la abstinencia. Para ambos experimentos, mantuvimos a las ratas en la instalación de animales bajo un ciclo inverso de luz / oscuridad de 12:12 horas con acceso libre a comida estándar de laboratorio y agua en sus jaulas domésticas durante todo el experimento. http://grants.nih.gov/grants/olaw/Guide-for-the-Care-and-Use-of-Laboratory-Animals.pdf). En el Experimento 1, 14 ratas completaron con éxito el entrenamiento de discriminación. Se excluyó una rata hembra debido a la falla de permeabilidad del catéter y una rata macho debido a la imposibilidad de adquirir la autoadministración del fármaco. Dos ratas macho y una rata hembra murieron durante el período de abstinencia. En el Experimento 2, las 16 ratas completaron con éxito el entrenamiento de discriminación. Una rata macho murió durante el período de abstinencia. Para ambos experimentos, utilizamos modelos multinivel basados en la máxima verosimilitud (SAS Proc Mixed) para dar cuenta de los datos faltantes.
Drogas
Recibimos 100 mg / ml de cocaína-HCl (cocaína) diluido en solución salina estéril de la farmacia NIDA. Elegimos una dosis unitaria de 0,75 mg / kg por infusión para el entrenamiento de autoadministración basado en estudios previos ( Koya et al., 2009 ) y mantuvimos la misma dosis unitaria durante el entrenamiento de discriminación.
Cirugía intravenosa
Para el Experimento 1, implantamos a las ratas con catéteres silásticos en su vena yugular derecha utilizando métodos previamente descritos ( Adhikary et al., 2017) Anestesiamos a las ratas con gas isofluorano (5% de inducción, 1-3% de mantenimiento) e insertamos catéteres silásticos en la vena yugular. Pasamos los catéteres por vía subcutánea a la región escapular media y los unimos a cánulas modificadas de calibre 22 (PlasticsOne, EE. UU.) Cementadas en malla de polipropileno (Small Parts Inc, EE. UU.) Colocadas debajo de la piel. Administramos ketoprofeno (2.5 mg / kg, inyección subcutánea; Henry Schein Inc, EE. UU.) Después de la cirugía para aliviar el dolor y permitimos que las ratas se recuperen durante 5 a 7 días antes del entrenamiento de autoadministración de drogas. Enjuagamos los catéteres diariamente con solución salina estéril que contenía gentamicina (4,25 mg / ml; Fresenius Kabi, EE. UU.) Durante las fases de recuperación y entrenamiento. Pesamos las ratas antes de cada sesión diaria de comportamiento, en el transcurso de cada experimento.
Aparato
Capacitamos y probamos a todas las ratas en cámaras de autoadministración estándar de Med Associates (Med Associates Inc, EE. UU.) (Med Associates ENV-007) encerradas en un gabinete ventilado y atenuador de sonido con ventanas oscurecidas. Cada cámara estaba equipada con un piso de rejilla de acero inoxidable y dos paredes laterales, cada una con tres paneles modulares operantes. Para el Experimento 1, equipamos la pared del lado derecho de la cámara con una sola palanca retráctil en el panel central, a 7,5 cm sobre el piso de la rejilla. Colocamos un estímulo discriminativo (luz, Med Associates ENV-221M) que señalaba la disponibilidad de cocaína en el panel izquierdo y otro estímulo discriminativo (luz, Med Associates ENV-221M) que señalaba la falta de disponibilidad de cocaína en el panel derecho de la misma pared lateral, equidistante de la palanca retráctil central y 14.0 cm sobre el piso de la rejilla. Además de la ubicación, utilizamos tapas de lentes rojas o blancas para diferenciar entre las dos señales discriminatorias y las contrarrestamos a través de las 14 cajas utilizadas para el Experimento 1. Conectamos el catéter de la rata a un dispositivo giratorio líquido (Instech Laboratories Inc, EE. UU.) A través de polietileno. 50 tubos que estaban protegidos por un resorte de metal y usaban una jeringa de 20 ml accionada por una bomba de jeringa de una sola velocidad (Med Associates PHM-100, 3.33 RPM) colocada fuera del gabinete de atenuación de sonido para administrar infusiones intravenosas de cocaína. En el Experimento 2, utilizamos ocho cámaras de autoadministración diferentes. Equipamos la pared lateral izquierda de estas cámaras con una sola palanca retráctil en el panel central, a 7,5 cm sobre el piso de la rejilla. Colocamos un estímulo discriminativo (luz, Med Associates ENV-221M) que señaló la disponibilidad de una recompensa de alimentos sabrosos en el panel derecho y otro estímulo discriminatorio (luz, Med Associates ENV-221M) que señaló la falta de disponibilidad de recompensas de alimentos en el panel izquierdo de la misma pared lateral, equidistante del palanca central retráctil y 14.0 cm sobre el piso de rejilla. Nuevamente usamos tapas de lentes rojas o blancas para diferenciar entre las dos señales discriminatorias y las contrarrestamos a través de las cajas utilizadas para este experimento. Equipamos el panel central de la pared opuesta (derecha) con un receptáculo de gránulos (Med Associates ENV-200R2M-6.0) conectado a un dispensador de gránulos de 45 mg (Med Associates ENV-203–45) para entregar una recompensa de comida sabrosa. equidistante de la palanca retráctil central y 14.0 cm sobre el piso de la rejilla. Nuevamente usamos tapas de lentes rojas o blancas para diferenciar entre las dos señales discriminatorias y las contrarrestamos a través de las cajas utilizadas para este experimento. Equipamos el panel central de la pared opuesta (derecha) con un receptáculo de gránulos (Med Associates ENV-200R2M-6.0) conectado a un dispensador de gránulos de 45 mg (Med Associates ENV-203–45) para entregar una recompensa de comida sabrosa. equidistante de la palanca retráctil central y 14.0 cm sobre el piso de la rejilla. Nuevamente usamos tapas de lentes rojas o blancas para diferenciar entre las dos señales discriminatorias y las contrarrestamos a través de las cajas utilizadas para este experimento. Equipamos el panel central de la pared opuesta (derecha) con un receptáculo de gránulos (Med Associates ENV-200R2M-6.0) conectado a un dispensador de gránulos de 45 mg (Med Associates ENV-203–45) para entregar una recompensa de comida sabrosa.
Procedimientos de comportamiento
Las líneas de tiempo experimentales para cada experimento se muestran en las Figuras 1A y 2A . Las fases de autoadministración, juicio y discriminación para los experimentos con cocaína y alimentos se describen por separado a continuación. Las posteriores fases de prueba de abstinencia y recaída son las mismas para ambos experimentos y se describen juntas
Autoadministración de cocaína
Entrenamos ratas machos y hembras para presionar la palanca para obtener recompensa de cocaína durante dos sesiones de 3 horas por día que se separaron por 30-60 min. Dimos a las ratas Froot Loops (Kellogg Company, EE. UU.) En su jaula local 1 día antes del inicio del entrenamiento y luego usamos Froot Loops triturados cuando fue necesario para alentar a las ratas a que se enganchen con la palanca durante el entrenamiento inicial de acceso continuo. El inicio de una sesión fue señalado por la iluminación de una señal de luz en el lado derecho de la palanca retráctil seguido 30 s más tarde por la presentación de la palanca retráctil central durante 180 minutos. La luz permaneció encendida durante la sesión y sirvió como un estímulo discriminatorio para la disponibilidad de recompensas de cocaína. La misma luz se utilizó más tarde como un estímulo discriminatorio para indicar la disponibilidad de cocaína durante el entrenamiento de discriminación basado en el juicio. A lo largo de la sesión, las respuestas en esta palanca fueron recompensadas bajo un esquema de refuerzo de proporción fija 1 (FR1) y la cocaína en una dosis unitaria de 0.75 mg / kg / infusión (0.1 ml / infusión) fue entregada durante 3.5 s. Esta duración de la infusión también sirvió como el período de tiempo de espera, durante el cual las presiones de palanca se registraron pero no se reforzaron. Es importante tener en cuenta que el suministro de cocaína no se combinó con ninguna señal discreta. Al final de cada sesión de 3 horas, se apagó el estímulo discriminativo y se retiró la palanca. Grabamos ( Es importante tener en cuenta que el suministro de cocaína no se combinó con ninguna señal discreta. Al final de cada sesión de 3 horas, se apagó el estímulo discriminativo y se retiró la palanca. Grabamos ( Es importante tener en cuenta que el suministro de cocaína no se combinó con ninguna señal discreta. Al final de cada sesión de 3 horas, se apagó el estímulo discriminativo y se retiró la palanca. Grabamos (O’Brien, 2005 ) el número total de prensas de palanca y ( Wikler, 1973 ) el número total de infusiones recibidas durante toda la sesión. Dimos a las ratas hasta seis sesiones de entrenamiento para adquirir una autoadministración estable que respondiera en el procedimiento de acceso continuo antes de cambiarlas a entrenamiento de prueba para obtener una recompensa de cocaína.
Entrenamiento de prueba para recompensa de cocaína
Entrenamos ratas en dos sesiones de entrenamiento de prueba de 3 horas por día durante 2 días. Dimos sesiones de entrenamiento de prueba de ratas antes del entrenamiento de discriminación basada en el juicio a ( O’Brien, 2005 ) habituar a las ratas al formato de prueba y ( Wikler, 1973 ) presentamos las dos posibles contingencias de prueba por separado antes de mezclarlas juntas durante las sesiones de discriminación. La figura 1B muestra la línea de tiempo para una prueba DS única, así como las diferencias entre los dos tipos de prueba.. Cada sesión en esta fase consistió en 30 ensayos discretos separados por un intervalo variable entre ensayos: el inicio de cada ensayo se señaló mediante la iluminación de un estímulo discriminatorio durante 30 s, después de lo cual las ratas tuvieron acceso a la palanca retráctil central durante 60 s. Durante este entrenamiento de prueba inicial, cada sesión consistió en solo uno de los dos tipos de prueba posibles: pruebas en las que la recompensa de cocaína estaba disponible (DS + ensayos) o pruebas en las que la recompensa de cocaína no estaba disponible (pruebas DS).
Los ensayos DS + fueron señalados por el mismo DS utilizado durante la autoadministración de acceso continuo (luz en el lado derecho de la palanca, contrarrestada para luz roja o blanca). Durante los ensayos DS +, las respuestas en la palanca fueron recompensadas bajo un esquema de refuerzo de proporción fija 1 (FR1) y la recompensa de cocaína a una dosis unitaria de 0.75 mg / kg / infusión (0.1 ml / infusión) se entregó durante 3.5 s. Esta duración de la infusión también sirvió como el período de tiempo de espera, durante el cual las presiones de palanca se registraron pero no se reforzaron. Las prensas de palanca adicionales durante este período de 60 s también se reforzaron en el mismo horario. Similar al entrenamiento de autoadministración, el suministro de cocaína en estos ensayos no se combinó con señales discretas. Sesenta segundos después de la presentación de la palanca, el DS + se apagó y la palanca se retrajo, señalando el final de la prueba.
Los ensayos DS se indicaron mediante el otro DS disponible (luz en el lado izquierdo de la palanca, contrarrestada para luz roja o blanca). Durante los ensayos DS, todas las respuestas en la palanca se registraron pero no se reforzaron. Sesenta segundos después de la presentación de la palanca, el DS- se apagó y la palanca se retrajo, señalando el final de la prueba.
Todas las ratas fueron entrenadas en ensayos DS + en la sesión de la mañana y ensayos DS en la sesión de la tarde. Utilizamos dos medidas de comportamiento para monitorear el entrenamiento durante esta fase: ( O’Brien, 2005 ) el número total de ensayos DS + vs. DS con al menos una palanca y ( Wikler, 1973 ) el número total de respuestas realizadas durante DS + vs DS-ensayos durante cada sesión de 3 horas.
Entrenamiento de discriminación por recompensa de cocaína
Capacitamos a las ratas en el procedimiento de discriminación basado en ensayos durante dos sesiones de 3 horas por día, separadas por al menos 30 minutos. En cada una de estas sesiones, las ratas recibieron un total de 60 ensayos discretos; 30 ensayos DS + y ensayos DS se entremezclaron y presentaron en un orden pseudoaleatorio de manera que las ratas no recibieron más de dos presentaciones consecutivas del mismo tipo de ensayo durante la sesión. Similar a la fase previa de entrenamiento, registramos ( O’Brien, 2005 ) el número total de ensayos DS + vs. DS- con al menos una palanca y ( Wikler, 1973 ) el número total de respuestas realizadas durante DS + vs. DS – ensayos durante cada sesión de 3 h.
Autoadministración de alimentos
Entrenamos ratas machos y hembras para presionar con palanca para obtener una recompensa de comida sabrosa (TestDiet, EE. UU.; Catálogo # 1811155, 12.7% de grasa, 66.7% de carbohidratos y 20.6% de proteína) durante una sesión de 1 hora por día. Dimos a las ratas los gránulos de comida de 45 mg en su jaula casera 1 día antes del comienzo del entrenamiento y luego usamos gránulos de comida triturados cuando era necesario para que las ratas se engancharan con la palanca durante el entrenamiento inicial de acceso continuo. El inicio de una sesión fue señalado por la iluminación de una señal luminosa a la derecha de una palanca retráctil central seguida 30 s más tarde por la presentación de la palanca retráctil durante 60 minutos. La luz permaneció encendida durante la sesión y sirvió como un estímulo discriminatorio para la disponibilidad de recompensas de alimentos sabrosos. La misma luz se usó luego como un estímulo discriminativo para indicar la disponibilidad de una recompensa de comida sabrosa durante el entrenamiento de discriminación basado en ensayos. A lo largo de la sesión, las respuestas en esta palanca fueron recompensadas bajo un cronograma de refuerzo de proporción fija 1 (FR1). La finalización exitosa del requisito de FR condujo a la entrega de tres gránulos de comida ‘preferidos’ o sabrosos de 45 mg durante 3.5 s. Esta duración de la entrega de recompensas no se combinó con ninguna señal discreta y sirvió como el período de tiempo de espera, durante el cual las presiones de palanca se registraron pero no se reforzaron. Al final de la sesión de 1 hora, se apagó el estímulo discriminativo y se retiró la palanca. Grabamos ( La finalización exitosa del requisito de FR condujo a la entrega de tres gránulos de comida ‘preferidos’ o sabrosos de 45 mg durante 3.5 s. Esta duración de la entrega de recompensas no se combinó con ninguna señal discreta y sirvió como el período de tiempo de espera, durante el cual las presiones de palanca se registraron pero no se reforzaron. Al final de la sesión de 1 hora, se apagó el estímulo discriminativo y se retiró la palanca. Grabamos ( La finalización exitosa del requisito de FR condujo a la entrega de tres gránulos de comida ‘preferidos’ o sabrosos de 45 mg durante 3.5 s. Esta duración de la entrega de recompensas no se combinó con ninguna señal discreta y sirvió como el período de tiempo de espera, durante el cual las presiones de palanca se registraron pero no se reforzaron. Al final de la sesión de 1 hora, se apagó el estímulo discriminativo y se retiró la palanca. Grabamos (O’Brien, 2005 ) el número total de prensas de palanca y ( Wikler, 1973 ) el número total de recompensas recibidas durante toda la sesión. Dimos a las ratas hasta tres sesiones de entrenamiento para adquirir una autoadministración estable que respondiera antes de cambiarlas a entrenamiento de prueba para obtener una recompensa de comida sabrosa.
Entrenamiento de prueba para la recompensa de comida sabrosa
Entrenamos ratas en dos sesiones de entrenamiento de prueba de 1 hora en 2 días. Dimos a las ratas dos sesiones de entrenamiento de prueba antes del entrenamiento de discriminación basada en el juicio para ( O’Brien, 2005 ) habituar a las ratas al formato de prueba y ( Wikler, 1973 ) presentamos las dos posibles contingencias de prueba por separado antes de mezclarlas juntas durante las sesiones de discriminación. La línea de tiempo para una única prueba de DS y las diferencias entre los dos tipos de prueba se muestran en la Figura 2B. Cada sesión en esta fase consistió en 30 ensayos discretos separados por un intervalo variable entre ensayos: el inicio de cada ensayo se señaló mediante la iluminación de un estímulo discriminatorio durante 30 s, después de lo cual las ratas tuvieron acceso a la palanca retráctil central durante 60 s. Durante esta capacitación inicial de prueba, cada sesión consistió en solo uno de los dos tipos de prueba posibles: pruebas en las que había una recompensa alimentaria aceptable (pruebas DS +) o pruebas en las que no había una recompensa alimentaria aceptable (pruebas DS).
Los ensayos DS + fueron señalados por el mismo DS utilizado durante la autoadministración de acceso continuo (luz en el lado derecho de la palanca, contrarrestada para luz roja o blanca). Durante los ensayos DS +, las respuestas en la palanca fueron recompensadas bajo un esquema de refuerzo de relación fija 1 (FR1). La finalización de FR dio como resultado la entrega de un único gránulo de comida sabrosa de 45 mg después de 1 sy un período de espera de 3,5 s durante el cual se registraron las presiones de palanca pero no se reforzaron. Las prensas de palanca adicionales durante este período de 60 s también se reforzaron en el mismo horario. Al igual que en el entrenamiento de autoadministración, la entrega de recompensa alimentaria en estos ensayos no se combinó con señales discretas. Sesenta segundos después de la presentación de la palanca, el DS + se apagó y la palanca se retrajo, señalando el final de la prueba.
Los ensayos DS se indicaron mediante el otro DS disponible (luz en el lado izquierdo de la palanca, contrarrestada para luz roja o blanca). Durante los ensayos DS, todas las respuestas en la palanca se registraron pero no se reforzaron. Sesenta segundos después de la presentación de la palanca, el DS- se apagó y la palanca se retrajo, señalando el final de la prueba.
Todas las ratas fueron entrenadas en ensayos DS + en la sesión de la mañana y ensayos DS en la sesión de la tarde. Utilizamos dos medidas de comportamiento para monitorear el entrenamiento durante esta fase: ( O’Brien, 2005 ) el número total de ensayos DS + vs. DS con al menos una palanca y ( Wikler, 1973 ) el número total de respuestas realizadas durante DS + vs DS-ensayos durante cada sesión de 3 horas.
Entrenamiento de discriminación para recompensas de comida sabrosa
Luego capacitamos a las ratas en el procedimiento de discriminación basado en el juicio para una sesión de 2 horas por día. En cada una de estas sesiones, las ratas recibieron un total de 60 ensayos discretos; 30 ensayos DS + y ensayos DS se entremezclaron y presentaron en un orden pseudoaleatorio de manera que las ratas no recibieron más de dos presentaciones consecutivas del mismo tipo de ensayo durante la sesión. Similar a la fase previa de entrenamiento, registramos ( O’Brien, 2005 ) el número total de ensayos DS + vs. DS- con al menos una palanca y ( Wikler, 1973 ) el número total de respuestas realizadas durante DS + vs. DS – ensayos durante cada sesión de 3 h.
Fase de abstinencia
Durante la fase de abstinencia para ambos experimentos, alojamos ratas en jaulas individuales en la instalación de animales y las manejamos 1 o 2 veces por semana. En el Experimento 1, después de que las ratas adquirieron con éxito la discriminación, las alojamos en el vivero por hasta 400 días adicionales y las probamos repetidamente para detectar recaídas después de períodos de abstinencia de cocaína cada vez más largos. En el Experimento 2, después de que las ratas adquirieron con éxito la discriminación, las alojamos en el vivero por hasta 200 días adicionales y las probamos repetidamente en busca de recaídas después de períodos de abstinencia progresivamente más largos de la recompensa de alimentos sabrosos.
Prueba de recaída
En los Experimentos 1 y 2, las condiciones experimentales durante las pruebas de recaída fueron las mismas que en la sesión de entrenamiento de discriminación basada en el ensayo correspondiente, excepto que las respuestas en la palanca no se reforzaron en los ensayos DS + o DS (condiciones de extinción). En el Experimento 1, las bombas de infusión se apagaron durante todas las pruebas de recaída y todas las ratas supervivientes se probaron 1, 21, 60, 120, 200, 300 y 400 días después de la última sesión de entrenamiento de discriminación. En el Experimento 2, los dispensadores de pellets se apagaron durante todas las pruebas de recaída y todas las ratas supervivientes se analizaron 1, 21, 60, 120 y 200 días después de la última sesión de entrenamiento de discriminación. Al igual que con el entrenamiento de discriminación, registramos ( O’Brien, 2005 ) el número total de ensayos DS + vs. DS- con al menos una palanca y ( Wikler, 1973) el número total de respuestas realizadas durante los ensayos DS + vs. DS- durante toda la sesión de prueba de recaída. Definimos operativamente el término recaída como la continuación de la búsqueda de drogas discriminada no reforzada después de un período de abstinencia.
Prueba de restablecimiento cebada con cocaína
En el Experimento 1, después de la prueba final de recaída (día 400), probamos las ratas (n = 11) para el restablecimiento inducido por cebado de cocaína durante cuatro sesiones separadas, ejecutadas en días consecutivos (días 401-404). En los días de prueba para el restablecimiento inducido por cebado de cocaína, les dimos a las ratas una inyección intraperitoneal (IP) de solución salina o cocaína 10 minutos antes del inicio de la sesión de prueba. Elegimos un orden de dosis ascendente para la cocaína en orden (10, 20 mg / kg) para minimizar el efecto de arrastre de una dosis de cebado dada en la dosis de cebado subsiguiente. Probamos las mismas ratas para el restablecimiento cebado con solución salina antes y entre las pruebas de restablecimiento cebado con cocaína. Las condiciones experimentales durante la prueba de reincorporación fueron las mismas que en la sesión de entrenamiento de discriminación basada en el juicio, excepto que las bombas de infusión se apagaron durante la duración de la prueba y las respuestas en la palanca no se reforzaron en los ensayos DS + o DS- (condiciones de extinción). Al igual que con el entrenamiento de discriminación y las pruebas de recaída, registramos (a) el número total de ensayos DS + vs. DS con al menos una palanca y (b) el número total de respuestas realizadas durante los ensayos DS + vs. DS durante toda la cocaína de sesión de prueba de reinicio preparada. Las dosis de cebado de cocaína se basaron en estudios previos utilizando el modelo de restablecimiento ( Pruebas de DS durante toda la sesión de prueba de restablecimiento cebada con cocaína. Las dosis de cebado de cocaína se basaron en estudios previos utilizando el modelo de restablecimiento ( Pruebas de DS durante toda la sesión de prueba de restablecimiento cebada con cocaína. Las dosis de cebado de cocaína se basaron en estudios previos utilizando el modelo de restablecimiento (Kalivas y McFarland, 2003 ).
Análisis estadístico
Como se describió anteriormente, no todas las ratas completaron todas las fases de los experimentos. En el Experimento 1, dos de las 16 ratas no completaron el entrenamiento de discriminación y fueron excluidas del estudio. Tres de las 14 ratas de prueba restantes murieron durante la abstinencia y no completaron todas las pruebas de recaída. En el Experimento 2, las 16 ratas completaron el entrenamiento de discriminación y fueron evaluadas repetidamente por recaída durante la abstinencia. Una rata de prueba murió durante la abstinencia y no completó todas las pruebas de recaída. Por lo tanto, utilizamos modelos multinivel basados en la máxima verosimilitud (SAS Proc Mixed) en lugar de análisis de varianza de medidas repetidas de mínimos cuadrados ordinarios para dar cuenta de los datos faltantes. Ambos enfoques logran los mismos objetivos, pero los modelos de máxima verosimilitud evitan la imputación de datos faltantes y permiten un modelado más preciso de la no homogeneidad de la varianza en puntos de tiempo desigualmente espaciados.
Llevamos a cabo todos los análisis estadísticos de dos medidas de comportamiento: ( O’Brien, 2005 ) el número total de ensayos de cada tipo de DS con al menos una palanca (denotada como ensayos «exitosos» ) y ( Wikler, 1973 ) el número total de respuestas realizadas durante cada tipo de prueba de DS durante toda la sesión (indicadas como prensas de palanca) Seguimos los principales efectos estadísticamente significativos o interacciones con pruebas post-hoc como se describe a continuación. Debido a que algunos de nuestros modelos produjeron múltiples efectos e interacciones principales, informamos solo aquellos que son críticos para la interpretación de datos. En los análisis preliminares que controlan el sexo, vimos diferencias de sexo en la adquisición de discriminación, pero no en los efectos de interés (por ejemplo, la intensidad de la búsqueda potenciada o el curso temporal de incubación de la respuesta controlada por DS). Por lo tanto, colapsamos nuestros análisis a través del sexo para ambos experimentos. Los datos desagregados por sexo para todas las fases de cada experimento se proporcionan como complementos de figuras vinculados a las figuras principales ( Figura 1 — figura suplemento 1 y Figura 2 — figura suplemento 1) y los resultados estadísticos para todos los análisis, incluidos los que controlan el sexo, se proporcionan como archivos de datos fuente asociados ( Figura 1: datos fuente 1 y Figura 2: datos fuente 1 ).
En el Experimento 1, para el análisis de la discriminación ( Figura 1C , n = 14), utilizamos un modelo factorial bidireccional con factores de sujetos dentro de la sesión de entrenamiento de discriminación (sesiones 5–14) y tipo de DS (DS +, DS-) , acompañado de la prueba de Tukey’s Honest Significant Difference (HSD) cuando sea apropiado para comparaciones por pares entre DS + y DS- para cada sesión de entrenamiento. Para las pruebas de recaída repetidas ( Figura 1D, n = 11–14), utilizamos modelos factoriales mixtos de dos vías con la duración de los factores dentro de los sujetos de la abstinencia forzada (1, 21, 60, 120, 200, 300 y 400 días) y el tipo de DS (DS +, DS -), seguido de la prueba de Dunnett para comparaciones por pares entre la prueba de recaída del día 1 y cada una de las pruebas de recaída de los días siguientes. También utilizamos HSD de Tukey para comparaciones por pares entre DS + y DS- para cada día de prueba de recaída.
Para las pruebas de restablecimiento inducido por cebado ( Figura 1E , n = 11), utilizamos modelos de 2 vías con los factores dentro de los sujetos dosis de cebado de cocaína (0, 10, 0, 20 mg / kg) y tipo de DS (DS +, DS-). Utilizamos el HSD de Tukey para las comparaciones por pares entre diferentes dosis de cebado de cocaína dentro de cada tipo de prueba de DS. También utilizamos el HSD de Tukey para comparaciones por pares entre DS + y DS- para cada día de prueba de reincorporación.
En el Experimento 2, para el análisis de la discriminación ( Figura 2C , n = 16), utilizamos un modelo factorial bidireccional con factores dentro de la materia de la sesión de entrenamiento de discriminación (sesiones 3-13) y tipo de DS (DS +, DS-) , acompañado de la prueba HSD de Tukey para comparaciones por pares entre DS + y DS- para cada sesión de entrenamiento. Para las pruebas de recaída repetidas ( Figura 2D , n = 15-16), utilizamos modelos factoriales de dos vías con la duración de los factores dentro de los sujetos de la abstinencia forzada (1, 21, 60, 120 y 200 días) y el tipo de DS ( DS +, DS-), seguido de las pruebas de Dunnett para comparaciones por pares entre la prueba de recaída del día 1 y cada una de las pruebas de recaída de los días siguientes. También utilizamos HSD de Tukey para comparaciones por pares entre DS + y DS- en cada día de prueba de recaída.
En todos los modelos, utilizamos una estructura de error de potencia espacial para dar cuenta de la autocorrelación en intervalos desigualmente espaciados; Esto es similar al uso de una corrección Huynh-Feldt o Greenhouse-Geisser en un ANOVA de medidas repetidas. El nivel alfa (significancia) se estableció en 0.05, dos colas.
Referencias
Incubation of nicotine seeking is associated with enhanced protein kinase A-regulated signaling of dopamine- and cAMP-regulated phosphoprotein of 32 kDa in the insular cortex. A AbdolahiG AcostaFJ BreslinSE HembyWJ Lynch (2010) European Journal of Neuroscience 31:733–741. https://doi.org/10.1111/j.1460-9568.2010.07114.x
Incubation of extinction responding and cue-induced reinstatement, but not context- or drug priming-induced reinstatement, after withdrawal from methamphetamine. S AdhikaryD CaprioliM VenniroP KallenbergerY ShahamJM Bossert (2017) Addiction Biology 22:977–990. https://doi.org/10.1111/adb.12386
Incubation of cue-induced cigarette craving during abstinence in human smokers. G BediKL PrestonDH EpsteinSJ HeishmanGF MarroneY ShahamH de Wit (2011) Biological Psychiatry 69:708–711. https://doi.org/10.1016/j.biopsych.2010.07.014
Time-dependent changes in alcohol-seeking behaviour during abstinence. P BienkowskiA RogowskiA KorkoszP MierzejewskiK RadwanskaL KaczmarekA Bogucka-BonikowskaW Kostowski (2004) European Neuropsychopharmacology 14:355–360. https://doi.org/10.1016/j.euroneuro.2003.10.005
Hierarchical and binary associations compete for behavioral control during instrumental biconditional discrimination. LA BradfieldBW Balleine (2013) Journal of Experimental Psychology: Animal Behavior Processes 39:2–13. https://doi.org/10.1037/a0030941
Stimuli associated with a single cocaine experience elicit long-lasting cocaine-seeking. R CiccocioppoR Martin-FardonF Weiss (2004) Nature Neuroscience 7:495–496. https://doi.org/10.1038/nn1219
Evidence for the hierarchical structure of instrumental learning. RM ColwillRA Rescorla (1990) Animal Learning & Behavior 18:71–82. https://doi.org/10.3758/BF03205241
The craving to smoke in flight attendants: relations with smoking deprivation, anticipation of smoking, and actual smoking. R DarN Rosen-KorakinO ShapiraY GottliebH Frenk (2010) Journal of Abnormal Psychology 119:248–253. https://doi.org/10.1037/a0017778
Associative theories of goal-directed behaviour: a case for animal-human translational models. S de WitA Dickinson (2009) Psychological Research Psychologische Forschung 73:463–476. https://doi.org/10.1007/s00426-009-0230-6
Circuit and synaptic plasticity mechanisms of drug relapse. Y DongJR TaylorME WolfY Shaham (2017) The Journal of Neuroscience 37:10867–10876.https://doi.org/10.1523/JNEUROSCI.1821-17.2017
Compulsive seekers: our take. two clinicians’ Perspective on a new animal model of addiction. DH EpsteinWJ Kowalczyk (2018) Neuropsychopharmacology 43:677–679. https://doi.org/10.1038/npp.2017.132
Haloperidol prevents the reinstatement of amphetamine-rewarded runway responding in rats. A Ettenberg (1990) Pharmacology Biochemistry and Behavior 36:635–638. https://doi.org/10.1016/0091-3057(90)90268-M
Persistent cue-evoked activity of accumbens neurons after prolonged abstinence from self-administered cocaine. UE GhitzaAT FabbricatoreV ProkopenkoAP PawlakMO West (2003) The Journal of Neuroscience 23:7239–7245. https://doi.org/10.1523/JNEUROSCI.23-19-07239.2003
Incubation of cocaine craving after withdrawal. JW GrimmBT HopeRA WiseY Shaham (2001) Nature 412:141–142. https://doi.org/10.1038/35084134
Effect of cocaine and sucrose withdrawal period on extinction behavior, cue-induced reinstatement, and protein levels of the dopamine transporter and tyrosine hydroxylase in limbic and cortical areas in rats. JW GrimmY ShahamBT Hope (2002) Behavioural Pharmacology 13:379–388. https://doi.org/10.1097/00008877-200209000-00011
Time-dependent increases in brain-derived neurotrophic factor protein levels within the mesolimbic dopamine system after withdrawal from cocaine: implications for incubation of cocaine craving. JW GrimmL LuT HayashiBT HopeTP SuY Shaham (2003) The Journal of Neuroscience 23:742–747. https://doi.org/10.1523/JNEUROSCI.23-03-00742.2003
The infralimbic and prelimbic cortices contribute to the inhibitory control of cocaine-seeking behavior during a discriminative stimulus task in rats. AL GutmanVA EwaldCV CosmeWR WorthRT LaLumiere (2017) Addiction Biology 22:1719–1730. https://doi.org/10.1111/adb.12434
Brain circuitry and the reinstatement of cocaine-seeking behavior. PW KalivasK McFarland (2003) Psychopharmacology 168:44–56. https://doi.org/10.1007/s00213-003-1393-2
Reinstatement of alcohol-seeking behavior by drug-associated discriminative stimuli after prolonged extinction in the rat. SN KatnerJG MagalongF Weiss (1999) Neuropsychopharmacology 20:471–479. https://doi.org/10.1016/S0893-133X(98)00084-0
Role of ventral medial prefrontal cortex in incubation of cocaine craving. E KoyaJL UejimaKA WihbeyJM BossertBT HopeY Shaham (2009) Neuropharmacology 56:177–185. https://doi.org/10.1016/j.neuropharm.2008.04.022
Incubation of methamphetamine and palatable food craving after punishment-induced abstinence. IN KrasnovaNJ MarchantB LadenheimMT McCoyLV PanlilioJM BossertY ShahamJL Cadet (2014) Neuropsychopharmacology 39:2008–2016. https://doi.org/10.1038/npp.2014.50
Incubation of alcohol craving during abstinence in patients with alcohol dependence. P LiP WuX XinYL FanGB WangF WangMY MaMM XueYX LuoFD YangYP BaoJ ShiHQ SunL Lu (2015a) Addiction Biology 20:513–522. https://doi.org/10.1111/adb.12140
Recent updates on incubation of drug craving: a mini-review. X LiD CaprioliNJ Marchant (2015b) Addiction Biology 20:872–876. https://doi.org/10.1111/adb.12205
Cocaine seeking over extended withdrawal periods in rats: different time courses of responding induced by cocaine cues versus cocaine priming over the first 6 months. L LuJW GrimmJ DempseyY Shaham (2004a) Psychopharmacology 176:101–108. https://doi.org/10.1007/s00213-004-1860-4
Recent developments in animal models of drug relapse. NJ MarchantX LiY Shaham (2013) Current Opinion in Neurobiology 23:675–683. https://doi.org/10.1016/j.conb.2013.01.003
Perseveration of craving: effects of stimuli conditioned to drugs of abuse versus conventional reinforcers differing in demand. R Martin-FardonF Weiss (2017) Addiction Biology 22:923–932. https://doi.org/10.1111/adb.12374
Reinstatement of drug-seeking behavior produced by heroin-predictive environmental stimuli. K McFarlandA Ettenberg (1997) Psychopharmacology 131:86–92. https://doi.org/10.1007/s002130050269
Discriminative inhibitory control of cocaine seeking involves the prelimbic prefrontal cortex. C MihindouK GuillemS NavaillesC VouillacSH Ahmed (2013) Biological Psychiatry 73:271–279. https://doi.org/10.1016/j.biopsych.2012.08.011
A review of preclinical research demonstrating that drug and Non-Drug ReinforcersDifferentially affect behavior. D N. KearnsM A. Gomez-SerranoB J. Tunstall (2011) Current Drug Abuse Reviewse 4:261–269. https://doi.org/10.2174/1874473711104040261
Fos protein expression and cocaine-seeking behavior in rats after exposure to a cocaine self-administration environment. JL NeisewanderDA BakerRA FuchsLT Tran-NguyenA PalmerJF Marshall (2000) The Journal of Neuroscience 20:798–805. https://doi.org/10.1523/JNEUROSCI.20-02-00798.2000
Classical conditioning in drug-dependent humans. CP O’BrienAR ChildressAT McLellanR Ehrman (1992) Annals of the New York Academy of Sciences 654:400–415. https://doi.org/10.1111/j.1749-6632.1992.tb25984.x
Anticraving medications for relapse prevention: a possible new class of psychoactive medications. CP O’Brien (2005) American Journal of Psychiatry 162:1423–1431. https://doi.org/10.1176/appi.ajp.162.8.1423
Incubation of Cue-Induced craving in adults addicted to cocaine measured by electroencephalography. MA ParvazSJ MoellerRZ Goldstein (2016) JAMA Psychiatry 73:1127–1134. https://doi.org/10.1001/jamapsychiatry.2016.2181
Neurobiology of the incubation of drug craving. CL PickensM AiravaaraF ThebergeS FanousBT HopeY Shaham (2011) Trends in Neurosciences 34:411–420. https://doi.org/10.1016/j.tins.2011.06.001
Diverse roads to relapse: a discriminative cue signaling cocaine availability is more effective in renewing cocaine seeking in goal trackers than sign trackers and depends on basal forebrain cholinergic activity. KK PitchersKB PhillipsJL JonesTE RobinsonM Sarter (2017) The Journal of Neuroscience 37:7198–7208. https://doi.org/10.1523/JNEUROSCI.0990-17.2017
Evidence for an association between the discriminative stimulus and the response-outcome association in instrumental learning. RA Rescorla (1990) Journal of Experimental Psychology: Animal Behavior Processes 16:326–334. https://doi.org/10.1037/0097-7403.16.4.326
Two-process learning theory: relationships between pavlovian conditioning and instrumental learning. RA RescorlaRL Solomon (1967) Psychological Review 74:151–182. https://doi.org/10.1037/h0024475
The role of neuroadaptations in relapse to drug seeking. Y ShahamBT Hope (2005) Nature Neuroscience 8:1437–1439. https://doi.org/10.1038/nn1105-1437
Time-dependent changes in extinction behavior and stress-induced reinstatement of drug seeking following withdrawal from heroin in rats. U ShalevM MoralesB HopeJ YapY Shaham (2001) Psychopharmacology 156:98–107. https://doi.org/10.1007/s002130100748
The anxiogenic drug yohimbine reinstates methamphetamine seeking in a rat model of drug relapse. JD ShepardJM BossertSY LiuY Shaham (2004) Biological Psychiatry 55:1082–1089. https://doi.org/10.1016/j.biopsych.2004.02.032
Cocaine can generate a stronger conditioned reinforcer than food despite being a weaker primary reinforcer. BJ TunstallDN Kearns (2016) Addiction Biology 21:282–293. https://doi.org/10.1111/adb.12195
Punishment models of addictive behavior. LJMJ VanderschurenAM MinnaardJAS SmeetsHMB Lesscher (2017) Current Opinion in Behavioral Sciences 13:77–84. https://doi.org/10.1016/j.cobeha.2016.10.007
Animal models of drug relapse and craving: from drug priming-induced reinstatement to incubation of craving after voluntary abstinence. M VenniroD CaprioliY Shaham (2016) Progress in Brain Research 224:25–52. https://doi.org/10.1016/bs.pbr.2015.08.004
Effects of length of abstinence on decision-making and craving in methamphetamine abusers. G WangJ ShiN ChenL XuJ LiP LiY SunL Lu (2013) PLOS ONE 8:e68791. https://doi.org/10.1371/journal.pone.0068791
Discriminated response and incentive processes in operant conditioning: a two-factor model of stimulus control. SJ Weiss (1978) Journal of the Experimental Analysis of Behavior 30:361–381. https://doi.org/10.1901/jeab.1978.30-361
Control of cocaine-seeking behavior by drug-associated stimuli in rats: effects on recovery of extinguished operant-responding and extracellular dopamine levels in amygdala and nucleus accumbens. F WeissCS Maldonado-VlaarLH ParsonsTM KerrDL SmithO Ben-Shahar (2000) PNAS 97:4321–4326. https://doi.org/10.1073/pnas.97.8.4321
Advances in the Neuroscience of Addiction. F Weiss (2010) CM Kuhn, GF Koob, editors. Boca Raton: CRC Press/Taylor & Francis.
The Instrumentally-Derived Incentive-Motivational function. SJ Weiss (2014) International Journal of Comparative Psychology, 27.
Synaptic mechanisms underlying persistent cocaine craving. ME Wolf (2016) Nature Reviews Neuroscience 17:351–365. https://doi.org/10.1038/nrn.2016.39
Basolateral amygdala lesions impair both cue- and cocaine-induced reinstatement in animals trained on a discriminative stimulus task. IA YunHL Fields (2003) Neuroscience 121:747–757. https://doi.org/10.1016/S0306-4522(03)00531-1
En el presente trabajo se revisaron los aspectos generales del craving y se destacó su importancia para la adherencia al tratamiento en adicciones. De igual manera se revisaron las diferentes estrategias cognitivo-conductuales para el manejo del craving, las cuales incluían: detención del pensamiento, técnicas de distracción, control y programación de actividades, entrenamiento en relajación, técnicas de imaginación, desensibilización sistemática, auto instrucciones y discusión de ideas irracionales. Esto se logró revisando la bibliografía pertinente. Se concluyó que cumplió con el objetivo de informar y contribuir al conocimiento del psicólogo.
Introducción
El manejo del craving es una situación determinante para la adherencia al tratamiento de las personas farmacodependientes, puesto que es el factor más importante de abandono terapéutico. Los pensamientos, sentimientos y conductas que se presentan durante el periodo de abstinencia pueden lograr, por su intensidad y su naturaleza, que el paciente deje el tratamiento y reincida dado el poder que éste ejerce sobre la persona, incluso, después de varios años de abstinencia. Por lo tanto, el craving significa un problema muy difícil para las personas que presentan abuso de sustancias. De igual manera no resta de importancia clínica.
El DSM IV, menciona que el craving se observa con frecuencia (quizá en todos) en los pacientes farmacodependientes, a la vez que describe como una “necesidad irresistible de consumo” (APA, 1994, pag. 182). Esa necesidad tiende a la realización de la misma, que se alcanza con la consumación por medio de un acto consumatorio. “Los craving y los impulsos por el consumo tienden a ser automáticos y llegar a ser «autónomos», es decir, pueden continuar incluso aunque la persona intente suprimirlos, no se suprimen con facilidad” (Sánchez 2002). Por lo anterior se destaca la relevancia de este documento, dado que su abordaje es de inminente importancia para el tratamiento del sujeto farmacodependiente, en donde el psicólogo debe conocer las estrategias que le competen para el mismo integrarse dentro de un equipo multidisciplinario y brindar un servicio de alta calidad al paciente para contribuir a su recuperación y rehabilitación. De esta manera, se pretende que el presente trabajo sirva como una guía sobre las estrategias para el manejo del craving, las cuales son, por elección, cognitivo conductuales, así como contribuir al conocimiento del lector. Para este efecto se revisarán los aspectos relevantes para este manejo.
El craving es un deseo (o necesidad psicológica) intenso e irrefrenable que conduce al individuo a abandonar la abstinencia. Beck y cols. (1999) identifican cuatro tipos de craving, que se enumeran a continuación:
Respuesta a los síntomas de la abstinencia: Cuando sucede la disminución del grado de satisfacción que encuentran en el uso de la droga, pero el incremento de sensación de malestar interno cuando deja de tomar la droga.
Respuesta a la falta de placer: Cuando sucede el aburrimiento o no tengan habilidades para encontrar formas prosociales de divertirse, por lo cual desea mejorar su estado de ánimo «automedicándose»
Respuesta «condicionada» a las señales asociadas con drogas: Cuando los pacientes asocian estímulos neutros (una esquina determinada de una calle, un número telefónico, un nombre, etc.) condicionados por una intensa recompensa obtenida de la utilización de drogas que inducen un craving automático.
Respuesta a los deseos hedónicos: Cuando el paciente combina una experiencia natural placentera con el uso de drogas (por ejemplo, cuando combina el sexo y drogas) que hace que se asocien, desarrollando un inicio de craving al realizar la actividad natural sin el uso de drogas por que el placer experimentado de estas experiencias no es comparable cuando se combina con la droga.
Es importante señalar que el manejo multidisciplinario profesional es vital para la efectividad del tratamiento. En función del manejo psicológico de las farmacodependencias, es decir, excluyendo las estrategias farmacológicas usadas en psiquiatría, las técnicas cognitivo conductuales son las de elección dado que se ha comprobado que son superiores a los otros métodos psicoterapéuticos.
“La literatura existente confirma la efectividad de las intervenciones de corte cognitivo-conductual en el tratamiento de las drogas en general combinados con el tratamiento médico.” (Sánchez y cols., 2002b, pag. 9). Ávila (2002) menciona que entre los tratamientos psicológicos más eficaces se encontraron como principales las terapias cognitivas. Así mismo encontró que las menos eficaces son las psicoanalíticas, psicoterapia general y terapia familiar. Otro beneficio de las estrategias cognitivo-conductuales, es que “con ellas el sujeto se siente mayormente comprometido con su abstinencia que con otras.” (Shine, 2000, pag. 1)
El terapeuta debe reconocer y comprender los cravings de su paciente, sus tipos, sus elementos esenciales y su naturaleza, así como empatizar con el mismo para poder determinar la mejor estrategia para este manejo. Para esto es relevante que el sujeto entienda el craving, para lo cual el paciente debe darse cuenta de que “experimentar cierto grado de ansiedad es normal y muy común. Sentir que esta ansiedad no significa que algo está mal o que el paciente realmente desea consumir drogas otra vez.” (NIDA, 1999, citado por CIJ, 1999, pag. 88) De igual manera, debe describir o detectar el craving. Para esto puede ser útil el empleo de autorregistros, así como preguntar ¿Cómo es su craving?, ¿Qué tanto le afecta?, ¿Cuánto tiempo dura? ¿Qué es lo que hace o trata de hacer para manejar el craving?, etc.
Ya identificado el craving y las emociones y pensamientos asociados (por medio de autorregistros, análisis funcional y la entrevista), se procede, según la elección del terapeuta y la naturaleza del craving, a la aplicación de un programa o estrategia cognitivo-conductual. A continuación, se describen algunos:
Detención de Pensamiento
El método de detención de pensamiento (También llamada bloqueo o parada de pensamiento) consiste en la interrupción y modificación, por medio de la sustitución, de los pensamientos asociados al craving. Es decir, “se entrena el paciente para que excluya, incluso antes de su formulación, cualquier pensamiento indeseable o improductivo.” (Wolpe, 1993, pag. 123) Esto conduce a la reducción de la cadena de pensamientos que se acompañan de emociones negativas, por lo tanto “las emociones negativas quedan cortadas antes de que puedan surgir” (Wolpe, 1969, citado por Sánchez, 2002a, pag. 39)
¿Cómo se aplica el método de detención de pensamiento?
Se procede a discutir con el paciente los pensamientos relacionados con el craving que son necesarios eliminar. Posterior a esto, el terapeuta le pide al paciente que cierre los ojos e inicie verbalizando con una secuencia de pensamientos negativos asociados al craving como, por ejemplo: «Me siento horrible, me voy a morir si no uso droga, etc.» Mientras el paciente procede, el terapeuta interrumpe diciendo «¡Alto!» Después de esto el terapeuta “hace ver al paciente cómo se detuvieron aquellos pensamientos” (Wolpe, 1993, pag. 124), pidiéndole que repita el ejercicio varias veces. Estos ejercicios se repiten con el propósito de que el paciente aprenda a detener estos pensamientos negativos gritando «¡Alto!» por sí mismo cuando estos se presenten, acción que el terapeuta deberá alentar y reforzar.
Puede ser que el paciente exprese que es difícil gritar «¡Alto!» si los pensamientos se presentan en un lugar público como un restaurante, una sala de cine u otro lugar de alta concurrencia social, algo que puede resultar embarazoso para el paciente. Para esto, se puede indicar al paciente que en los lugares que considere pertinentes no realice la verbalización «¡Alto!», pero que puede pensarlo de manera firme y decidida, de modo que el paciente esté consciente de la manera en la que detuvo el pensamiento. Al terminar la detención del pensamiento, existe la adición de sustitución posterior del pensamiento negativo por uno incompatible o distractorio. Por ejemplo, en vez de “me voy a morir si no uso drogas”, se sustituye por “estoy bien” o cualquier otro pensamiento positivo y útil para el sujeto.
Técnicas de Distracción
Como su nombre indica, son técnicas sencillas que tienen como objetivo «distraer» al sujeto ante pensamientos y sentimientos relacionados con el craving. Es decir, que “cambien su enfoque atencional de interno a externo, para que disminuyan sus deseos” (Sánchez, 2002a, pag. 39) La distracción puede consistir en que el terapeuta indique, con un previo acuerdo en función de las actividades de distracción de relevancia y agrado para el paciente, lo siguiente: centrar la atención en los elementos del ambiente verbalizándolos, siendo lo más detallista posible (por ejemplo, “en este restaurante hay diez mesas con manteles blancos, las paredes son azul claro con una ventana cada cuatro mesas. Hay una señora que…); evitar el ambiente (por ejemplo, alejarse de un bar, esquina o lugar de consumo. Puede ir a visitar un amigo, a dar un paseo, etc.); hablar sobre algún tema iniciando una conversación con amigo o familiar, recitar un poema o cantar una canción favorita. (Generalmente, algunos pacientes prefieren traerlo escrito y leerlo); realizar actividades domésticas, ejercicio (por ejemplo, salir a correr, realizar determinado número de sentadillas, etc.) jugar un videojuego, juego de mesa o rompecabezas. En general estas distracciones se pueden combinar y complementar según las particularidades del paciente y de su craving. Así mismo, es importante evaluar la necesidad de combinar una estrategia de distracción con un ejercicio de relajación para reducir la ansiedad en casos necesarios.
Control y Programación de Actividades
Es una técnica que tiene como objetivo modificar los hábitos del paciente en función de sus actividades, que por lo general giran en torno al uso de drogas. Así mismo, “cuando los pacientes se están recuperando deben enfrentarse con mucho tiempo libre en sus manos.” (Beck y cols., 1999, pag. 227) Por lo cual, es importante que las actividades previas a la abstinencia «relevantes al uso de droga» sean sustituidas por nuevas actividades positivas para el paciente.
Se procede inicialmente elaborando un registro línea base de las conductas relacionadas con el consumo del paciente. Se buscará establecer las actividades que fueron o que son de disfrute para el paciente de modo que pueda programar sus actividades el mismo, lo cual establecerá “fuentes de refuerzo no relacionadas con las drogas” (Beck y cols., 1999, pag. 228).
Entrenamiento en Relajación
El Entrenamiento en relajación son procedimientos que están enfocados a hacer incompatible estado de relajación con la ansiedad o la ira, emociones que pueden propiciar los cravings, por medio del entrenamiento del paciente de modo que el mismo propicie un estado de relajación al enfrentar situaciones de ansiedad o ira para él. “De manera típica un terapeuta entrena a un cliente para relajarse profundamente, para lo cual se encuentran disponibles muchos métodos para lograr la relajación.” (Kazdin, 1996, pag. 275)
¿Cuál es la técnica principalmente usada?
Wolpe (1993) afirma que “el método de relajación que se emplea de manera rutinaria es esencialmente el de Jacobson [1983], pero la instrucción se completa en seis lecciones, en contraste con el entrenamiento prolongado que él utilizaba. (generalmente 50 sesiones o más)” Primero, se le debe explicar al paciente acerca de la naturaleza de los ejercicios y su objetivo principal, que es una habilidad que se le va a enseñar y que requiere práctica, así como enfatizar el deseo del terapeuta de que el paciente lo practique en casa por lo menos de 10 a 15 minutos. Posterior a esto, se realizan varios ejercicios en las sesiones de tensión y distensión de los músculos. Se puede iniciar relajando los brazos; se le pide al paciente que tome el bracero de la silla o sillón para distinguir alguna diferencia (si es que existe) entre las sensaciones que se producen en su brazo y en su mano. Después, se sigue con la cabeza, ya que “por lo general, los efectos más marcados de inhibición de la ansiedad se obtienen relajando esa parte del cuerpo.” (Wolpe, 1993, pag. 154). De ahí se prosigue hacia abajo. En la segunda sesión, se le dice al paciente que se va a continuar con la relajación de los músculos de la cabeza, dado que estos son los más importantes desde el punto de vista emocional. Es importante señalarle al paciente la manera en la que se está trabajando de manera que se retroalimente. En la tercera sesión, se trabaja con los músculos de la boca y de la lengua. En la cuarta sesión, se prosigue con los músculos de la región del ojo. En la quinta sesión, se sigue con los músculos del cuello y los hombros. En la sexta sesión, se trata con los músculos de la espalda, del abdomen y del tórax. En la séptima sesión, se puede iniciar con los pies y hacia arriba finalmente. Es importante evaluar la reducción de los niveles de ansiedad así como de los cravings por medio de instrumentos de medición pertinentes para determinar en qué medida los ejercicios de relajación están ayudando al paciente.
Técnicas de Imaginación
Son técnicas en las cuales el paciente modifica sus pensamientos utilizando la imaginación. Estas técnicas pueden combinarse efectivamente con otras técnicas para incrementar las probabilidades de éxito en otros aspectos del tratamiento. Por ejemplo, “puede utilizarse la imaginación para que los pacientes visualicen el autocontrol y evitar el consumo de drogas. Es útil como método para cambiar las creencias relacionadas con las drogas y los pensamientos automáticos, o bien distrayéndolos de los craving y urgencias. También puede ser útil como método para cambiar las creencias y los pensamientos relacionados con las drogas.” (Sánchez, 2002a, pag. 40)
Beck y cols. (1999), destacan cinco técnicas utilizadas: a) reenfoque con imaginación, b) sustitución por imagen negativa, c) sustitución por imagen positiva, d) repaso mediante imagen, y e) adquirir dominio mediante imaginación.
Esta técnica sirve como complemento de la técnica de detención de pensamiento. Cuando el paciente empieza a presentar pensamientos negativos referentes al craving y dice «¡Alto!», puede imaginar una señal de alto (como las que uno encontraría en la calle), un semáforo en rojo o cualquier imagen que para él represente la acción de «¡Alto!», de modo que acentúa la detención. Después puede optar por emplear una de las técnicas de distracción (como, por ejemplo, centrar la atención en el ambiente o recitar un poema) de modo que pase el craving.
Sustitución por imagen negativa
Esta técnica se refiere a la imaginación de una imagen negativa acerca de las consecuencias negativas de consumir drogas cuando aparecen los pensamientos que conllevan al craving. De tal forma que las imágenes “positivas” acerca de la droga (por ejemplo, recordar la última vez que consumió) sea sustituida por una imagen acerca de las consecuencias negativas que tiene el último (p. e., el recuerdo de una hospitalización debida al consumo o los dolores físicos de la abstinencia) para evitar la conducta de uso.
Reemplazo por imágenes positivas
Se refiere a la sustitución de imágenes negativas referentes al consumo de drogas relacionadas con las creencias irracionales como las predicciones negativas (por ejemplo, creer que el uso de drogas ha arruinado su vida para siempre y terminará viviendo en la calle), y que se puede sustituir por una “tarjeta flash”, que describa una situación positiva. Todo esto con el objetivo de disminuir las emociones negativas y consecuentemente, el craving.
Repaso mediante la imagen
En ésta, las imágenes se utilizan como medio de “práctica” o repaso de ciertas habilidades adquiridas (por ejemplo, habilidades sociales o resolución de problemas) o de la desensibilización ante situaciones o ambientes que pueden ser disparadores de craving (por ejemplo, una fiesta en donde le ofrezcan una bebida). Por ejemplo “Un paciente que se había abstenido a la cocaína y el alcohol durante un año, planeó ir a un banquete formal donde se servía alcohol. En el repaso mediante a la imaginación el terapeuta instó al paciente a imaginar que iba al banquete y que decía cuando se le ofrecía una bebida: «No, gracias, tomaré agua.»”
Dominio mediante la imaginación
Esto implica la repetición y la gestación de una autoimagen poderosa y fuerte, para poder superar los cravings en su imaginación y para poder proyectarla hacia la realidad posteriormente. Puede utilizarse la metáfora para obtener poder ante el craving (por ejemplo, el paciente puede imaginar que él es un beisbolista y que el craving es la pelota que él batea y se va lejos).
Desensibilización sistemática
Es una técnica que emplea la exposición real o imaginaria a los estímulos que producen el craving, por medio de acercamientos sistemáticos a estos, los cuales son reforzados puntualmente; de modo que la respuesta pierda poder ante dicha situación dado que “la repetida presentación del estímulo hace que éste pierda progresivamente su capacidad de evocar ansiedad.” (Wolpe, 1993, pag. 145) Generalmente, se inicia un repaso mediante la imaginación del acercamiento al estímulo condicionado y, después de haberlo dominado, se prosigue la exposición al estímulo real.
Auto instrucciones y Autoafirmaciones
Se refiere a la verbalización de palabras que sugieren al paciente qué es lo que debe hacer o sentir, realizadas por el mismo sujeto. “El entrenamiento en auto instrucción se ha utilizado como una técnica de modificación conductual, pues con ella se entrena a los individuos para que hagan sugerencias y comentarios específicos que guíen su propia conducta de modo parecido a ser instruidos por alguien más.” (Kazdin, 1996, pag. 307) Por ejemplo, «Me siento bien, el craving va a pasar», «No necesito drogas para ser feliz», «¡Vas Bien!», «Recuerda que con lo que te ahorras en droga, ahora puedes comprarte la ropa que te gusta». En este sentido, también se pueden utilizar las tarjetas flash, las cuales son frases de afrontamiento al craving escritas en tarjetas de cartulina de un tamaño tal que el individuo pueda cargar con ellas a donde vaya. “Como ejemplo, éstas son algunas de las frases que un paciente escribió en sus tarjetas flash: 1. Te sientes más sano cuando no tomas drogas; 2. Las cosas me van muy bien con mi mujer; ¡sigue así!; 3. Tienes buen aspecto físico; ¡sigue así!; 4. ¡Vete de aquí inmediatamente!
Discusión de ideas irracionales
Técnica basada en la terapia racional emotiva (Ellis y Driden, 1987) en donde se “tratan de discutir de manera activa y directa todas las creencias irracionales que llevan a los pacientes a perturbarse a sí mismos por su adicción” (Ellis y Driden, citado por Sánchez, 2002a, pag. 40) Es una estrategia cognoscitiva que debate las distorsiones cognoscitivas del paciente, y que se pueden resumir en: extrapolar (por ejemplo: “o todo o nada”, “no debo sentir nada de craving por que si no significa que soy un perdedor), “adivinar” el pensamiento de otros (por ejemplo: “como caí en las drogas, todos piensan que soy un tonto”, “ese señor que está ahí me está viendo porque sabe que yo usaba drogas”), predicciones negativas (por ejemplo: “nunca voy a cambiar”, “siempre voy a estar sintiéndome mal si no uso la droga”, “me voy a encontrar en la calle a todos mis amigos que usan droga y voy a recaer”), exacerbar lo negativo (por ejemplo: “todo me sale mal, no hay nada bueno en mi vida”, “¡me siento horrible, si no uso la droga, me voy a morir!”), negligencia y descalificación ante lo positivo (por ejemplo: “si me felicitó, fue por puro compromiso, él ha de pensar que voy a volver a usar droga”), minimización de lo positivo y amplificación de lo negativo (por ejemplo: “que haya dejado la droga es bueno, pero lo que hice es imperdonable e inolvidable”), sentimiento de estafa y perfeccionismo (por ejemplo: “debí haber controlado mi último craving, y como no lo hice, he decepcionado a todos, soy un fraude”).
Para combatir las ideas irracionales se debe confrontar y discutir las mismas, por ejemplo: “menciona que se siente horrible y que va a morir. Sin duda es molesto e indeseable el sentir el craving, pero, ¿en realidad se va usted a morir por no tomarte un trago en esa fiesta, sufre usted de alguna enfermedad extraña en la cual al no ingerir alcohol usted de repente cae súbitamente al suelo y deja de vivir? –el paciente ríe y responde— No, tal vez estoy exagerando.” De esta manera el paciente comprende la irracionalidad de sus creencias y cómo ha aprendido a percibir las cosas de una manera
inapropiada lo cual le ha generado emociones que no puede controlar. Por lo tanto, es necesario que adopte una nueva forma de pensamiento, el cual será eminentemente racional. “El pensamiento racional presenta un contraste acentuado respecto a su ilógico opuesto: Está basado en la realidad, contribuye a alcanzar nuestras metas y propósitos, crea emociones que podemos controlar; y nos ayuda a comportarnos de la manera que nos facilita nuestras aspiraciones y supervivencia…” (Froggatt, 1993, adaptado por Ureva, 2003)
Conclusión
El manejo del craving es determinante para el tratamiento psicológico de las adicciones y para la adherencia al tratamiento en general. El manejo de este dependerá de la elección del terapeuta entre la amplia gama de estrategias a utilizar y de su habilidad para manejarlas. Es esencial que previo al manejo de estas estrategias, se determine la naturaleza y tipo de craving que el paciente presenta para hacer la elección adecuada en función de las necesidades del paciente y de sus características y recursos personales.
Las diferentes técnicas han demostrado ser ampliamente efectivas en el tratamiento de otras patologías específicas relacionadas con la ansiedad. Por tanto, dada la naturaleza del craving, es ampliamente factible la efectividad de estas estrategias en el manejo del mismo. Se observa también, que, en muchos de los casos, dichas estrategias no están aisladas, sino que en ocasiones se complementan para ser más eficaces, por lo cual se sugiere adoptar una visión más amplia, en vez de pretender hacer uso de una sola.
Es evidente, que en este documento falta información y mayor explicación acerca de la aplicación de las estrategias cognitivo conductuales mencionadas. No obstante se cree que el objetivo de servir como guía y referencia clara de estas técnicas, de tal modo que el lector, después de conocer las generalidades de las mismas, busque información más precisa para aplicarlas efectivamente. De igual manera se cree que este ensayo contribuyó al aprendizaje para el psicólogo en función del conocimiento del trabajo que se hace en adicciones, específicamente, del manejo del craving.
La Bibliografía y las tablas de los artículos las pueden pedir a:
La pérdida de control sobre el consumo de drogas que se produce en la adicción se creía inicialmente como resultado de la interrupción de los circuitos subcorticales de recompensa. Sin embargo, los estudios de imagen en conductas adictivas han identificado una participación clave de la corteza prefrontal (CPF) tanto a través de su regulación de las regiones de recompensa límbica como su participación en la función ejecutiva de orden superior (por ejemplo, autocontrol, atribución de saliencia y conciencia). Esta revisión se centra en los estudios funcionales de neuroimagen realizados en la última década que han ampliado nuestra comprensión de la participación del CPF en la adicción a las drogas. La interrupción del CPF en la adicción subyace no solo al consumo compulsivo de drogas, sino también a las conductas desventajosas asociadas con la adicción y la erosión del libre albedrío.
La adicción a las drogas abarca un ciclo recurrente de intoxicación, atracones, abstinencia y deseo que produce un consumo excesivo de drogas a pesar de las consecuencias adversas (FIG. 1). Las drogas que son abusadas por los humanos aumentan la dopamina en el circuito de recompensa y se cree que esto subyace a sus efectos gratificantes. Por lo tanto, la mayoría de los estudios clínicos sobre la adicción se han centrado en las áreas de la dopamina del cerebro medio (el área tegmental ventral y la sustancia negra) y las estructuras de los ganglios basales a las que se proyectan (el estriado ventral, donde se encuentra el núcleo accumbens y el estriado dorsal), que se sabe están involucrados en la recompensa, el condicionamiento y la formación de hábitos 1 – 3. Sin embargo, estudios clínicos y preclínicos han sacado a la luz más recientemente y comenzaron a aclarar el papel de la corteza prefrontal (CPF) en la adicción 4 . Se atribuyen una serie de procesos al CPF que son fundamentales para una función neuropsicológica saludable (que abarca la emoción, la cognición y el comportamiento) y que ayudan a explicar por qué la alteración del CPF en la adicción podría afectar negativamente a una amplia gama de comportamientos (TABLA 1).
tabla 1 Procesos asociados con la corteza prefrontal que se interrumpen en la adicción.
Procesos
Posible perturbación en la adicción
Región probable de CPF
Autocontrol y monitoreo del comportamiento: inhibición de la respuesta, coordinación del comportamiento, predicción de conflictos y errores, detección y resolución.
Impulsividad, compulsividad, asunción de riesgos y autocontrol deteriorado (patrones de comportamiento habituales, automáticos, impulsados por estímulos e inflexibles)
DLCPF, dACC, IFG y vlCPF
Regulación de la emoción: supresión cognitiva y afectiva de la emoción.
Mayor reactividad al estrés e incapacidad para suprimir la intensidad emocional (por ejemplo, ansiedad y afecto negativo)
mOFC, vmCPF y subgenual ACC
Motivación: impulso, iniciativa, persistencia y esfuerzo hacia la consecución de objetivos.
Mayor motivación para adquirir drogas, pero menor motivación para otros objetivos y compromiso y esfuerzo comprometidos
OFC, ACC, vmCPF y DLCPF
Conciencia e interocepción: sentir el propio estado corporal y subjetivo, la percepción
Reducción de la saciedad, la «negación» de la enfermedad o la necesidad de tratamiento, y el pensamiento orientado externamente
rACC y dACC, mCPF, OFC y vlCPF
La atención y flexibilidad: la formación de fijar y de mantenimiento en comparación con la configuración de desplazamiento, y la tarea de conmutación
Sesgo de atención hacia estímulos relacionados con las drogas y lejos de otros estímulos y refuerzos, e inflexibilidad en los objetivos para obtener la droga.
DLCPF, ACC, IFG y vlCPF
Memoria de trabajo: memoria a corto plazo que permite construir representaciones y orientar la acción.
Formación de la memoria que está sesgada hacia estímulos relacionados con las drogas y lejos de las alternativas.
DLCPF
Aprendizaje y memoria: aprendizaje asociativo estímulo-respuesta, aprendizaje inverso, extinción, devaluación de recompensa, inhibición latente (supresión de información) y memoria a largo plazo
Condicionamiento farmacológico y capacidad alterada para actualizar el valor de recompensa de los refuerzos no farmacológicos
DLCPF, OFC y ACC
Toma de decisiones: valoración (refuerzos de codificación) versus elección, resultado esperado, estimación de probabilidad, planificación y formación de objetivos
La anticipación relacionada con las drogas, la elección de la recompensa inmediata sobre la gratificación retrasada, el descuento de las consecuencias futuras y las predicciones inexactas o la planificación de la acción
IOFC, mOFC, vmCPF y DLCPF
Atribución de saliencia: valoración del valor afectivo, saliente de incentivo y utilidad subjetiva (resultados alternativos)
Las drogas y las señales de las drogas tienen un valor sensibilizado, los reforzadores no farmacológicos se devalúan y los gradientes no se perciben, y el error de predicción negativa (la experiencia real es peor de lo esperado)
mOFC y vmCPF
La corteza orbitofrontal (OFC) incluye el área de Brodmann (BA) 10–14 y 47 ( REF. 216 ), y las regiones inferior y subgenual de la corteza cingulada anterior (ACC) (BA 24, 25 y 32) en la corteza prefrontal ventromedial (vmCPF) 217 ; El ACC incluye el ACC rostral (rACC) y el ACC dorsal (dACC) (BA 24 y 32, respectivamente), que se incluyen dentro del CPF medial (mCPF). El mCPF también incluye BA 6, 8, 9 y 10 (REF. 218); CPF dorsolateral (DLCPF) incluye BA 6, 8, 9 y 46 (REF. 219); y el giro frontal inferior (IFG) y el CPF ventrolateral (vlCPF) abarcan porciones inferiores de BA 8, 44 y 45 (REF. 220). Estos diversos procesos y regiones participan en un grado diferente en el deseo, intoxicación, atracones y abstinencia. IFC, lateral OFC; mOFC, OFC medial; CPF, corteza prefrontal.
Sobre la base de los hallazgos de imágenes y los estudios preclínicos emergentes 5 , 6 , propusimos hace 10 años que la función alterada de la CPF conduce a un síndrome de inhibición de la respuesta alterada y atribución de saliencia (iRISA) en la adicción (Figura 1), un síndrome que se caracteriza por atribuir una atención excesiva a las señales relacionadas con las drogas y los medicamentos, la sensibilidad disminuida a los reforzadores no farmacológicos y la capacidad reducida para inhibir las conductas desventajosas o inadaptadas 7 . Como resultado de estos déficits principales, la búsqueda y el consumo de drogas se convierten en un impulso motivacional principal, que se produce a expensas de otras actividades 8 y culmina en conductas extremas para obtener las drogas 9 .
Aquí revisamos los estudios de imagen sobre el papel del CPF en la adicción de la última década, integrándolos en el modelo iRISA con el objetivo de obtener una mayor comprensión de la disfunción del CPF en la adicción. Específicamente, esta es la primera evaluación sistemática del papel de distintas regiones dentro del CPF funcionalmente heterogéneo en los mecanismos neuropsicológicos que supuestamente subyacen al ciclo de recaída de la adicción. Revisamos los estudios de tomografía por emisión de positrones (TEP) y MRI funcional (fMRI, por sus siglas en inglés) que se centran en las regiones del CPF que se han relacionado con la adicción. Estos incluyen la corteza orbitofrontal (OFC), la corteza cingulada anterior (ACC) y la corteza prefrontal dorsolateral (DLCPF) (consulte la Tabla 1 para las áreas de Brodmann; consulte Información complementaria S1 (tabla) para las áreas de Brodmann que no se tratan en el texto principal). Consideramos los resultados de estos estudios (Figura 2) en el contexto del papel que desempeña el CPF en iRISA: primero, en la respuesta a los efectos directos de la droga y las señales relacionadas con la droga; segundo, en la respuesta a las recompensas no relacionadas con las drogas, como el dinero; tercero, en la función ejecutiva de orden superior, incluido el control inhibitorio; y cuarto, en conciencia de la enfermedad. Presentamos un modelo simple que ayuda a guiar nuestras hipótesis con respecto al papel de las distintas subregiones de CPF en el endofenotipo de la adicción a las drogas (Figura 3), como se describe con más detalle a continuación. Para los estudios preclínicos sobre el CPF en adicciones o cuentas en profundidad sobre la función ejecutiva del CPF, remitimos al lector a otras revisiones 10 ,11 .
Figura 2 Estudios recientes de neuroimagen de la actividad de CPF en individuos adictos a las drogas Las áreas de activación (medido mediante RM, tomografía por emisión de positrones (TEP) o tomografía computarizada por emisión de fotón único (SPECT)) ( Información complementaria S1 (tabla) ) se representan en un espacio estereotáxico, representado en las superficies dorsal y ventral (arriba) parte) y las superficies lateral y medial (parte media e inferior, respectivamente) del cerebro humano. una| Cambios de actividad relacionados con las características neuropsicológicas en la adicción. Las áreas de la corteza prefrontal (CPF) muestran diferencias en la actividad entre individuos con adicción y controles saludables durante las tareas que involucran atención y memoria de trabajo (mostradas en verde), toma de decisiones (mostradas en azul claro), control inhibitorio (mostradas en amarillo), emoción y motivación (mostrado en rojo), y reactividad de cue y administración del fármaco (mostrado en naranja). Además, en algunas áreas de CPF, la actividad se correlaciona con el desempeño de la tarea o el uso de drogas (se muestra en azul oscuro). segundo| Cambios en la actividad relacionados con las características clínicas de la adicción, que incluyen intoxicación y atracones (se muestran en rojo; los medicamentos se usaron dentro de las 48 horas posteriores al estudio), el deseo (se muestra en rosa, los medicamentos se utilizaron 1–2 semanas antes del estudio) y el retiro (se muestra en púrpura; las drogas se usaron más de 3 semanas antes del estudio). También se indican las áreas que mostraron activación en estudios en los que la etapa de adicción no se especificó o no se pudo determinar (se muestra en marrón). Estos son los mismos estudios que los descritos en a . Los estudios se incluyeron solo si se proporcionaron las coordenadas x , y y z, y si estas coordenadas estaban dentro de la materia gris de CPF; estudios en los que x , y y zLas coordenadas no se pudieron ubicar o se etiquetaron incorrectamente y no se incluyeron. Todas las coordenadas x , y y z se convirtieron al espacio Talairach (utilizando GingerAle, una aplicación Java multiplataforma para el metanálisis) antes de trazar. Se utilizó la caja de herramientas 213 , 214 del análisis de densidad de kernel multinivel (consulte el sitio web del software CANLab de la Universidad de Colorado).
figura 3 Un modelo de participación de CPF en iRISA en adicciones. Un modelo de cómo las interacciones entre las subregiones de la corteza prefrontal (CPF) pueden regular los cambios cognitivos, emocionales y de comportamiento en la adicción. El modelo muestra cómo los cambios en la actividad de las subregiones de CPF en individuos adictos se relacionan con los síntomas clínicos centrales de la adicción (intoxicación y atracones, y abstinencia y deseo) en comparación con la actividad de CPF en individuos o estados sanos y no adictos. El modelo se centra especialmente en el control inhibitorio y la regulación de la emoción. Los óvalos azules representan subregiones de CPF dorsal (incluido el CPF dorsolateral (DLCPF), la corteza cingulada anterior dorsal (dACC) y el giro frontal inferior; consulte la TABLA 1) que están involucrados en el control de orden superior (procesos ‘fríos’). Los óvalos rojos representan subregiones de CPF ventral (la corteza orbitofrontal medial (mOFC), CPF ventromedial y ACC rostroventral) que participan en procesos más automáticos relacionados con las emociones (procesos «calientes»). Las funciones neuropsicológicas relacionadas con las drogas (por ejemplo, la importancia del incentivo, la falta de drogas, el sesgo de atención y la búsqueda de drogas) que están reguladas por estas subregiones están representadas por tonos más oscuros y las funciones no relacionadas con las drogas (por ejemplo, esfuerzo sostenido) están representadas por tonos más claros. a) En el estado saludable, predominan las funciones cognitivas no relacionadas con las drogas, las emociones y los comportamientos (mostrados por los grandes óvalos de color claro) y las respuestas automáticas (las emociones y las tendencias de acción que podrían conducir al consumo de drogas) se suprimen mediante aportes del CPF dorsal (mostrado por la flecha gruesa). Por lo tanto, si una persona en estado saludable está expuesta a las drogas, se evita o detiene la conducta excesiva o inapropiada de consumo de drogas (‘Stop’). b) Durante el deseo y la abstinencia, las funciones cognitivas, las emociones y los comportamientos relacionados con las drogas comienzan a eclipsar las funciones no relacionadas con las drogas, creando un conflicto relacionado con el consumo de drogas («¿Detener?»). La disminución de la atención y / o el valor se asigna a estímulos no relacionados con las drogas (que se muestran en los óvalos más claros), y esta reducción se asocia con un autocontrol reducido y con la anhedonia, la reactividad al estrés y la ansiedad. También hay un aumento (mostrado por los óvalos oscuros más grandes) en la cognición sesgada por el fármaco y el deseo y deseo de drogas inducidos por la señal. c) Durante la intoxicación y el atracón, las funciones cognitivas de orden superior no relacionadas con las drogas (mostradas por el pequeño óvalo azul claro) se suprimen mediante la entrada incrementada (mostrada por la flecha gruesa) de las regiones que regulan las funciones «calientes» relacionadas con las drogas (grandes óvalos rojo oscuro). Es decir, hay una entrada disminuida de las áreas de control cognitivo de orden superior (que se muestra con una flecha punteada delgada), y las regiones «calientes» llegan a dominar la entrada cognitiva de orden superior. Por lo tanto, la atención se reduce para centrarse en las señales relacionadas con las drogas sobre todos los demás refuerzos, aumenta la impulsividad y se desatan las emociones básicas, como el miedo, la ira o el amor, según el contexto y las predisposiciones individuales. El resultado es que predominan los comportamientos automáticos impulsados por estímulos, como el consumo compulsivo de drogas, la agresión y la promiscuidad («¡Adelante!»). Este modelo no tiene en cuenta el desafío de localizar las funciones de PFC o la evidencia de que algunas personas adictas usan drogas para ‘automedicarse’ en un intento de normalizar las funciones de PFC (aunque la parte a podría representar una aproximación de las funciones de PFC normalizadas en estos individuos).
Al evaluar esta Revisión, los lectores deben aceptar una gran cantidad de resultados, lo que puede resultar bastante confuso ya que no siempre se proporcionan conclusiones definitivas. Esto es particularmente cierto para la localización de funciones: por ejemplo, ¿el dorsal ACC y el DLCPF están involucrados en la respuesta del deseo o en el control del deseo, o en ambos? La determinación de qué subregión de CPF media qué función puede ser muy difícil, presumiblemente debido a la flexibilidad neuroanatómica y cognitiva de estas funciones, es decir, los participantes pueden utilizar múltiples estrategias al realizar tareas neuropsicológicas, y los sistemas prefrontales parecen tener un mayor nivel de flexibilidad funcional que Más sistemas sensoriomotores primarios. Otra década de investigación puede resultar invaluable en nuestra comprensión del papel del CPF en la adicción a las drogas. RECUADRO 1 para una discusión de los primeros estudios que examinan estas redes en el contexto de la adicción, y el uso de modelos computacionales puede ayudar aún más a la hora de atribuir las probables funciones psicológicas a seleccionar las regiones de CPF y mejorar nuestra comprensión de su participación en la adicción a las drogas. Nuestra revisión es un paso en esta dirección.
Efectos directos de la exposición a drogas
Aquí, revisamos estudios que evaluaron los efectos de los fármacos estimulantes y no estimulantes sobre la actividad de CPF (información complementaria S2 (tabla). Nuestro modelo predice mejoras de la actividad inducidas por el fármaco en las áreas de CPF que están involucradas en los procesos relacionados con el fármaco, incluidas las respuestas emocionales, los comportamientos automáticos y la participación de ejecutivos de orden superior (por ejemplo, OFC medial (mOFC) y CPF ventromedial en el ansia, OFC en expectativa de drogas, ACC en sesgo de atención y DLCPF en la formación de memorias de trabajo relacionadas con las drogas). También predice disminuciones inducidas por el fármaco en la actividad no relacionada con el fármaco en estas mismas regiones CPF, especialmente durante el deseo y atracones en individuos adictos al fármaco, que se analizan a continuación (Figura 3). De acuerdo con la predicción anterior, la administración de cocaína por vía intravenosa a individuos adictos a la cocaína que se abstuvieron durante la noche aumentaron los autoinformes de alta y ansia, y principalmente aumentaron las respuestas dependientes del nivel de oxígeno en la sangre por RMN (BOLD, por sus siglas en inglés) en varias subregiones CPF 12, 13. Curiosamente, la actividad en la lateral lateral izquierda, la corteza frontopolar y el CAC se moduló por la expectativa de la droga (es decir, la actividad fue mayor después de la administración intravenosa de cocaína esperada frente a la inesperada), mientras que las regiones subcorticales respondieron principalmente a los efectos farmacológicos de la cocaína (es decir, no hubo modulación por expectativa); la dirección específica del efecto difería según la región de interés (ROI) 13 . En un 18El estudio de fluorodioxiglucosa PET (PET FDG), la administración del medicamento estimulante metilfenidato (MPH) a los usuarios activos de cocaína incrementó el metabolismo de la glucosa en todo el cerebro 14. En este caso, la COS lateral izquierda mostró un metabolismo mayor en respuesta al MPH inesperado que al esperado; el patrón opuesto al del efecto BOLD en el estudio anterior 13 posiblemente refleja la diferente sensibilidad temporal de las modalidades de imagen (ver más abajo).
Las drogas estimulantes también aumentan la actividad de CPF en animales de laboratorio. Por ejemplo, el flujo sanguíneo cerebral regional (rCBF) en monos rhesus sin uso de drogas aumentó en DLCPF después de la administración no contingente y en ACC durante una autoadministración simple de cocaína de tasa fija 15, 16. Un estudio de PET FDG en el mismo modelo animal mostró que la autoadministración de cocaína aumentaba el metabolismo en la OFC y el ACC en mayor medida cuando se extendía el acceso a la cocaína que cuando el acceso era limitado 17 (tenga en cuenta que el acceso extendido, pero no el acceso limitado o corto) se asocia con la transición de la ingesta de drogas moderada a excesiva, como ocurre en la adicción 18). De manera similar, la administración intracerebroventricular de cocaína en ratas indujo una gran respuesta de IRMf en regiones del cerebro seleccionadas, incluyendo CPF 19.
En conjunto, el efecto principal de la cocaína (y otros estimulantes como el MPH) en el CPF es aumentar la actividad de CPF, medida por el metabolismo de la glucosa, CBF o BOLD (aunque en un estudio reciente, la cocaína redujo el volumen de sangre cerebral de CPF en monos macacos). 20 A medida que la duración del acceso al fármaco y la expectativa de fármaco modulan la actividad de CPF, los aumentos en la actividad que se producen durante la administración del fármaco pueden ser indicativos de las adaptaciones neuroplásticas que se producen en la transición del primer uso u ocasional al uso regular, de manera tal los procesos, que incluyen la anticipación relacionada con las drogas (y otras respuestas condicionadas), suprimen o eclipsan los procesos no relacionados con las drogas, como la anticipación de, o la motivación para, perseguir objetivos no relacionados con las drogas ( FIG. 3 ).
En los fumadores de cigarrillos, el rCBF se redujo en el ACC dorsal izquierdo (dACC) y esto se correlacionó con una disminución del deseo después de fumar el primer cigarrillo del día 21. Se notificaron correlaciones similares entre rCBF en OFC y deseo después de inyecciones agudas de heroína en personas que dependen de la heroína 22. La disparidad entre los efectos de la cocaína (y otros estimulantes) y otros tipos de drogas en la actividad de CPF puede reflejar diferencias en los efectos farmacológicos directos de las drogas en la CPF y otras regiones del cerebro (cannabinoides, opioides mu y receptores de nicotina, que son los objetivos para la marihuana, la heroína y la nicotina, respectivamente, tienen una distribución regional distinta del cerebro) o en objetivos que no son del SNC (la cocaína y la metanfetamina tienen efectos simpaticomiméticos periféricos que son distintos de los efectos periféricos de la marihuana o el alcohol), o puede reflejar una variabilidad metodológica factores (por ejemplo, si los estudios analizaron valores absolutos o relativos (o normalizados)) 23. También puede estar relacionado con los efectos del deseo inducido por las drogas: con drogas como la cocaína, el deseo en las personas adictas aumenta de 10 a 15 minutos después de fumar, mientras que los estudios analizados informaron una disminución en el deseo inmediatamente después de la administración de nicotina o heroína. Visto de esta manera, y de acuerdo con nuestro modelo, los resultados colectivos sugieren que cuando la ingesta de medicamentos disminuye la ansiedad, esto se asocia con disminuciones en la actividad de CPF relacionada con las drogas, y viceversa. Concomitantemente con estas disminuciones relacionadas con las drogas, esperaríamos que aumentara la actividad de CPF no relacionada con las drogas, como es el caso (ver más abajo). Las disparidades entre los resultados en esta sección, y a lo largo de esta Revisión, también podrían atribuirse a las diferencias entre las diversas modalidades de obtención de imágenes, un problema que debe reconocerse al principio de esta Revisión. Por ejemplo, la PET FDG mide la actividad metabólica de la glucosa con un promedio de más de 30 minutos, mientras que fMRI BOLD y PET CBF reflejan cambios más rápidos en los patrones de activación. Estas modalidades también difieren en sus medidas de línea de base: no es posible establecer una línea de base absoluta con la RMF BOLD, mientras que es posible con la RM de PET y marcaje de giro arterial. Otra diferencia común entre los estudios es el estado de referencia de un individuo, por ejemplo, la duración de la abstinencia podría afectar las medidas de ansiedad y abstinencia.
Respuestas a las señales relacionadas con las drogas
En el núcleo de la adicción a las drogas se encuentran las respuestas condicionadas a los estímulos asociados con la droga que se desarrollan en los usuarios habituales, como los objetos que se utilizan para administrar la droga, las personas que adquieren la droga o los estados emocionales que en el pasado fueron liberados o activados. por el uso de la droga, que luego impulsa el deseo de tomar drogas y que son contribuyentes importantes para la recaída. Los estudios de imágenes han evaluado estas respuestas condicionadas al exponer a personas adictas a señales relacionadas con las drogas, por ejemplo, mostrándoles imágenes relacionadas con las drogas. Aquí, primero revisamos los estudios que compararon la respuesta de CPF con la exposición de referencia en individuos y controles adictos (Información complementaria S3 (tabla), y luego discutimos los estudios que exploraron el efecto de la abstinencia, la expectativa y las intervenciones cognitivas en las respuestas de CPF a las señales relacionadas con las drogas (Información complementaria S4 (tabla). Predecimos que, en individuos adictos, las respuestas de CPF a las señales relacionadas con las drogas imitan las respuestas a la droga en sí misma, debido al condicionamiento, y que la intervención causa una reducción de las respuestas condicionadas de la indicación de la droga en la CPF.
Efecto de la exposición de las señales sobre la actividad de CPF
Aunque hay algunas excepciones 24 – 26, los estudios de IRMf reportan que, en comparación con los controles, los individuos adictos a las drogas muestran una respuesta BOLD mejorada en CPF a las señales relacionadas con las drogas en relación con las señales de control (información complementaria S3 (tabla).
Estos resultados se informaron en DLCPF izquierdo, giro frontal medial izquierdo y giro subcalloso derecho (área de Brodmann 34) en fumadores jóvenes de cigarrillos 27, y en DLCPF y CAC bilaterales en alcohólicos abstinentes a corto plazo 28 y a largo plazo 29. Se notificaron aumentos similares en estudios (incluidos los estudios de PET con FDG) de personas adictas a la cocaína que miraban videos relacionados con la cocaína 30 y de grandes fumadores que miraban videos relacionados con el cigarrillo mientras manejaban un cigarrillo 31. A menudo, no hay diferencias entre los individuos adictos y no adictos en la valencia o en las calificaciones de excitación, o incluso en las reacciones autonómicas (por ejemplo, respuestas de conductancia de la piel) a las señales relacionadas con las drogas 29, lo que sugiere que las medidas de neuroimagen son más sensibles para detectar diferencias grupales en las respuestas condicionadas a señales relacionadas con las drogas. Es importante destacar que las respuestas de CPF inducidas por señales se correlacionaron con el deseo 31 y la severidad del uso de drogas 27, y predijeron el desempeño subsiguiente en una tarea de reconocimiento de emociones cebada 32 y el uso de drogas 3 meses después 29, lo que indica que estas medidas tienen relevancia clínica. Como no se activó CPF mediante señales enmascaradas relacionadas con el fármaco 33 (que activaron regiones subcorticales en lugar de 34), estos efectos solo pueden inducirse cuando se perciben conscientemente señales relacionadas con el fármaco, pero esto debe estudiarse más a fondo.
Una interesante línea de estudios explora la activación de CPF relacionada con la señal durante la exposición farmacológica aguda a drogas. En los machos dependientes de heroína que recibieron inyecciones de heroína mientras miraban videos relacionados con drogas, CBF en OFC se correlacionó con la necesidad de usar la droga y CBF en DLCPF (área 9 de Brodmann) se correlacionó con felicidad 22 (Información complementaria S2 (tabla). En este contexto, es interesante observar que el mero sabor del alcohol (en comparación con el jugo de litchi) puede aumentar la actividad de BOLD CPF en los bebedores jóvenes, y esta respuesta se correlaciona con el consumo de alcohol y el antojo de35 y posiblemente esté impulsada por la neurotransmisión de dopamina en la recompensa subcortical circuito 36. Por el contrario, en los bebedores de alcohol o fumadores de cigarrillos no dependientes, la actividad de la OFP relacionada con la señal se redujo mediante la administración de alcohol o nicotina, respectivamente [37]. Este hallazgo resuena con el hallazgo de que, en sujetos no adictos, la administración intravenosa de MPH disminuyó el metabolismo en las regiones ventrales de CPF 38 (RECUADRO 2). Los estudios futuros podrían comparar directamente las respuestas de CPF a las señales relacionadas con las drogas en individuos no dependientes y dependientes y, por lo tanto, explorar más a fondo el impacto de la intoxicación en las respuestas de CPF relacionadas con la señal. El modelado de atracones en sujetos que abusan de las drogas sería informativo para el diseño de intervenciones para reducir los comportamientos compulsivos inducidos por la señal.
Cuadro 2 El papel de la dopamina y otros neurotransmisores
Los receptores de dopamina D2, que se expresan más densamente en regiones subcorticales como el mesencéfalo y el estriado dorsal y ventral, también se distribuyen a lo largo de la corteza prefrontal (CPF). Una serie de estudios de tomografía por emisión de positrones (TEP) informaron una menor disponibilidad del receptor D2 de la dopamina estriatal en individuos adictos a la metanfetamina 184, cocaína 38 o alcohol 185, y en personas con obesidad mórbida 186 y estas reducciones se asociaron con una disminución de la actividad metabólica basal en la corteza orbitofrontal (OFC) y la corteza cingulada anterior (ACC). Esto sugiere que la pérdida de la señalización de la dopamina a través de los receptores D2 puede subyacer a algunos de los déficits en la función prefrontal que se observan en la adicción, una idea que está respaldada por datos preliminares que muestran que la disponibilidad del receptor D2 de la dopamina estriatal se correlacionó con la respuesta del CPF medial al dinero en la cocaína Individuos -Addicidos 187. La disponibilidad reducida del receptor D2 de la dopamina estriatal también se informó en fumadores masculinos, tanto después de fumar como de costumbre y después de 24 horas de abstinencia; en la condición de saciedad, la disponibilidad del receptor D2 de dopamina en el ACC bilateral se correlacionó negativamente con el deseo de fumar (se observaron correlaciones positivas para el cuerpo estriado y la OFC) 188. La evidencia de agotamiento de la dopamina en el dorsolateral CPF (córtex prefrontal dorsolateral) también se informó en los usuarios de ketamina crónicas jóvenes, y los niveles de agotamiento se correlaciona con el consumo de drogas semanal más alta 189. Otros estudios de PET informaron una liberación de dopamina estriatal notablemente atenuada en respuesta a la administración intravenosa de un fármaco estimulante (por ejemplo, metilfenidato) en consumidores de alcohol y alcohólicos, con una disminución paralela en las experiencias autonotificadas de sentirse alto 38, 185.
De acuerdo con los datos de estudios en animales, estos resultados en individuos adictos apuntan a una función dopaminérgica del estriado romo, tanto en la línea de base como en respuesta a un desafío directo, que se asocia con mayor deseo y severidad de uso. Una respuesta de dopamina estriatal embotada predice la elección real de la cocaína en lugar del dinero en individuos abstinentes adictos a la cocaína, lo que sugiere que puede predisponer a los sujetos a la recaída 190. Los resultados también sugieren que, al regular la magnitud de los aumentos de dopamina en el cuerpo estriado 185, la OFC asume un papel crucial en la modulación del valor de los reforzadores; la interrupción de esta regulación puede subyacer al aumento del valor atribuido a una recompensa de drogas en sujetos adictos. De acuerdo con esta sugerencia, el metabolismo en la OFC medial y el ACC ventral en los consumidores de cocaína aumentó después de la administración de estimulantes intravenosos, mientras que se redujo en los controles; los aumentos metabólicos regionales en los abusadores se asociaron con el deseo de drogas 38.
Los opioides endógenos también median las respuestas gratificantes de muchas drogas de abuso, en particular la heroína, el alcohol y la nicotina. El uso repetido de drogas se ha asociado con una disminución en la liberación de opioides endógenos, un efecto que puede contribuir a los síntomas de abstinencia, incluida la disforia. Un estudio que utilizó [11 C] carfentanilo demostró que los consumidores de cocaína tenían un mayor potencial de unión al receptor de opiáceos CPF mu (indicativo de niveles opiáceos endógenos más bajos) que los controles sanos no adictos, y que esto persistió en la corteza frontal anterior y el CAC durante las 12 semanas de abstinencia 191. El aumento de la unión del receptor opiáceo mu en el DLCPF y el ACC antes del tratamiento se asoció con un mayor uso de cocaína y una menor duración de la abstinencia, y se sugirió que era un mejor predictor del resultado del tratamiento que el consumo inicial de drogas y alcohol 192. Se informaron resultados similares en hombres alcohólicos abstinentes 193, mientras que la metadona crónica revierte el nivel de la unión del receptor opiáceo mu (o kappa) en individuos adictos a la heroína 194.
Se ha reportado una disminución del potencial de unión de CPF para un radioligando transportador de serotonina en los usuarios abstinentes de metanfetamina 195, los jóvenes usuarios recreativos de MDMA 196 y en los alcohólicos recuperados 197. La disponibilidad reducida del transportador de serotonina puede reflejar neuroadaptaciones al aumento de la serotonina sináptica, pero también podría reflejar el daño a las terminales nerviosas serotonérgicas. Otros sistemas de neurotransmisores que regulan el CPF y están involucrados en las neuroadaptaciones que ocurren con el uso repetido de drogas en animales de laboratorio incluyen el glutamato 198 y los sistemas de cannabinoides 199, 200. Sin embargo, hasta el momento no hay estudios publicados con radiotrazadores para visualizar estos sistemas en la adicción humana.
Consulte la información complementaria S7 (tabla) para obtener una descripción general de los estudios que comparan los sistemas de neurotransmisores entre individuos adictos y controles sanos.
La activación de CPF a señales relevantes también se ha informado en adicciones de comportamiento. Por ejemplo, los varones jóvenes que jugaron juegos por Internet durante más de 30 horas a la semana mostraron activaciones BOLD en OFC, ACC, CPF medial y DLCPF al ver las imágenes del juego, y estas activaciones se correlacionaron con la necesidad de jugar 39. De manera similar, en comparación con los sujetos de control, los jugadores patológicos que vieron videos de juegos de apuestas mostraron una mayor activación en la DLCPF derecha y el giro frontal inferior 40, y esta activación se correlacionó con la necesidad de apostar 41. Por el contrario, otro estudio en jugadores patológicos mostró una reducción de las respuestas de CPF BOLD ventromedial izquierdo para ganar en lugar de perder en una tarea similar al juego, y el tamaño de la reducción se correlacionó con la gravedad de la adicción al juego, según se evaluó con un cuestionario de juego 42. Las direcciones opuestas de los cambios de actividad (hiperactivaciones frente a hipoactivaciones en comparación con los controles) pueden ser impulsadas por el ROI (por ejemplo, las desactivaciones relacionadas con la tarea CPF ventromedial se ven a menudo y se han atribuido al papel de la red de «cerebro predeterminado» 43 ), diferencias en el deseo (el deseo se informó en REFS 39 – 41 pero no en el REF. 42), diferencias de tareas o factores metodológicos, que se resumen al final de esta sección.
Los trastornos que se caracterizan por un control deficiente del consumo de alimentos también se asocian con una reactividad anormal de CPF a las señales. Esto no es inesperado, dado que estos trastornos y la adicción implican compromisos similares en los circuitos neuronales 44, incluida la disminución de la disponibilidad del receptor D2 de dopamina estriatal 45. Por ejemplo, las mujeres con anorexia o bulimia que están viendo imágenes de alimentos de forma pasiva (en comparación con las imágenes no relacionadas con alimentos) mostraron un aumento de las respuestas de la prueba de IRMF en el CPF ventromedial izquierdo 46. En comparación con los pacientes con bulimia, los pacientes con anorexia mostraron una mayor activación de la OFC derecha en respuesta a las imágenes de los alimentos, lo que posiblemente implique a esta región en un autocontrol demasiado restrictivo; por el contrario, la actividad de la DLCPF izquierda a estas imágenes disminuyó en pacientes con bulimia en comparación con los controles sanos, lo que posiblemente implique a esta región en la pérdida de control sobre la ingesta de alimentos 46. En otro estudio, las mujeres jóvenes con trastornos de la alimentación, pero no sujetos de control, mostraron activación del CPF ventromedial izquierdo durante la selección de la palabra más negativa de los conjuntos de palabras relacionadas con la imagen corporal negativa (en comparación con durante la selección de la palabra más neutral de conjuntos de palabras neutrales) 47. Tales diferencias no se observaron para las palabras generalmente negativas, lo que indica que la activación de esta región se debió a las palabras que están más fuertemente relacionadas con las preocupaciones reales de este grupo de pacientes. Tomados junto con los resultados en los jugadores patológicos descritos anteriormente 42 , las respuestas ventromediales de CPF pueden rastrear la relevancia emocional de las señales de mayor preocupación para la población de pacientes en cuestión (es decir, ganar o evitar la pérdida para individuos con juego patológico, imagen corporal para individuos con trastornos de la alimentación y señales relacionadas con las drogas para los individuos adictos a las drogas) y podría servir como un objetivo para el seguimiento de las intervenciones terapéuticas en la adicción, como se sugirió recientemente 48 , 49 .
Efecto de la abstinencia, expectativa e intervenciones cognitivas
Aquí, proponemos que la intervención cognitiva y la abstinencia a largo plazo atenúan las respuestas inducidas por señales en el CPF, y que la expectativa relacionada con las drogas y la abstinencia a corto plazo tienen el efecto opuesto. El impacto de la abstinencia a corto plazo en la actividad relacionada con la señal de CPF se ha estudiado más ampliamente en la adicción a la nicotina (información complementaria S4 (tabla). En un estudio de IRM con rotaciones arteriales, la abstinencia de 12 horas en los fumadores aumentó el deseo, la CBF global y la CBF regional en la OFC, y la CBF disminuyó en la CPF derecha, con cambios de la CBF en todas las ROI que se correlacionan con los síntomas de la ansiedad y la abstinencia 50. Dicha reactividad de señal aumentada también se informó durante períodos más prolongados de abstinencia: hasta 8 días en DLCPF, ACC y giro frontal inferior en mujeres fumadoras 51– y también correlaciona positivamente con el deseo 52. Sin embargo, algunos estudios informan que la abstinencia no tiene efecto en la actividad de CPF inducida por el estímulo 53. Esto podría posiblemente atribuirse a otros factores que contribuyen a la variabilidad sustancial de los resultados, como la expectativa de fumar al final del estudio 54. De hecho, como se discutió anteriormente 13, solo la expectativa puede imitar los efectos de la ingesta aguda de medicamentos en la activación de CPF en individuos adictos. Los estudios en los que se exploran las tres variables (la expectativa de administración de drogas, la exposición a estímulos relacionadas con las drogas y la abstinencia) se analizan para determinar los efectos principales y los efectos de interacción en la actividad de CPF serían útiles, especialmente si involucran muestras grandes. La dinámica temporal de la reactividad de la señal de CPF también debe explorarse en estudios longitudinales, rastreando al mismo individuo durante períodos de abstinencia a más largo plazo.
Una línea de investigación prometedora explora la modulación del comportamiento de la reactividad de la señal. Por ejemplo, un hallazgo de un estudio reciente de PET en consumidores de cocaína sugirió un papel para la mOFC en la supresión del deseo de fumar. El deseo de fumar aumentó después de ver un video de las señales relacionadas con el coco, y los niveles de deseo se correlacionaron con el metabolismo de la glucosa en el CPF medial 55. Es importante destacar que cuando los participantes recibieron instrucciones, antes de ver el video, de inhibir el deseo, el metabolismo en el mOFC derecho disminuyó, y esto se asoció con la activación del giro frontal inferior derecho (área de Brodmann 44), que es una región crucial en el control inhibitorio. En los fumadores de cigarrillos que buscan tratamiento, la instrucción de resistir el deseo cuando se ven videos relacionados con el hábito de fumar se asoció con la activación de DLCPF y ACC, aunque inesperadamente, esta activación se correlacionó positivamente con el deseo 56. Un estudio reciente sugiere que la dirección del cambio en la actividad y la correlación con el deseo puede ser modulada por la estrategia de comportamiento que se utiliza para suprimir el deseo. En este elegante estudio, se instruyó a los fumadores de cigarrillos para que consideraran las consecuencias inmediatas versus las consecuencias a largo plazo de consumir los estímulos descritos en imágenes (señales relacionadas con los cigarrillos y relacionadas con los alimentos) 57. Teniendo en cuenta que las consecuencias a largo plazo se asociaron con un aumento de la actividad en las regiones de CPF asociadas con el control cognitivo (DLCPF y giro frontal inferior) y con una menor actividad en las regiones de CPF asociadas con el deseo (mOFC y ACC). Además, el deseo de autoinforme disminuyó cuando los sujetos consideraron las consecuencias a largo plazo, y se correlacionó negativamente con la actividad en dACC y DLCPF. Un análisis de mediación mostró que la asociación entre el aumento de la actividad en la DLCPF y las disminuciones relacionadas con la regulación en el deseo ya no era significativa después de incluir una disminución de la actividad en el estriado ventral en el modelo. Sin embargo, los estudios preclínicos que utilizan herramientas de ablación u optogenética son necesarios para comprender mejor la interacción de la CPF y el cuerpo estriado ventral para suprimir las respuestas de deseo. Tomados en conjunto, HIGO. 3 ), que distingue entre las regiones CPF que facilitan el esfuerzo cognitivo no relacionado con las drogas y el control inhibitorio (DLCPF, dACC y giro frontal inferior) y aquellas que reflejan la preocupación emocional relacionada con las drogas, el deseo y las conductas compulsivas (mOFC y ACC ventral).
En resumen, la exposición a estímulos relacionados con las drogas imita los efectos de la administración directa de drogas sobre la actividad de CPF en individuos adictos a las drogas, aunque el impacto de la duración de la abstinencia y la expectativa de consumo de drogas (y procesos relacionados como la formación de memorias relacionadas con las drogas), y sus contribuciones únicas a la función de CPF, aún no se han evaluado en muestras de gran tamaño. Al ampliar los estudios de reactividad de referencia para incluir funciones neuropsicológicas adicionales, y al explorar la dirección de las correlaciones entre la actividad de CPF y los puntos finales específicos (por ejemplo, ansia), la importancia funcional de las activaciones de las regiones específicas de CPF en la adicción se hará más clara. Una recomendación adicional para futuros estudios sobre la reactividad del estímulo es realizar comparaciones directas entre sesiones (por ejemplo, abstinencia versus saciedad) y condiciones de la tarea (por ejemplo, señales de drogas versus neutras) y realizar correlaciones de todo el cerebro con los cambios de comportamiento respectivos. Los estudios futuros también podrían comparar la duración y el patrón de activación de CPF después de la exposición aguda al fármaco y después de la exposición a señales condicionadas en los mismos sujetos. Los estudios en individuos no adictos se podrían usar para evaluar el impacto de la privación (por ejemplo, de los alimentos) y las necesidades urgentes (por ejemplo, el hambre, el deseo sexual y la motivación para el logro) en la reactividad de las señales de CPF. Por ejemplo, en controles sanos jóvenes, el deseo de alimentos imaginados, inducido por una dieta monótona, se asoció con la activación en varias regiones límbicas y paralímbicas, incluida la ACC (área de Brodmann 24) señales de drogas frente a señales neutrales) y para realizar correlaciones de todo el cerebro con los cambios de comportamiento respectivos. Los estudios futuros también podrían comparar la duración y el patrón de activación de CPF después de la exposición aguda al fármaco y después de la exposición a señales condicionadas en los mismos sujetos. Los estudios en individuos no adictos se podrían usar para evaluar el impacto de la privación (por ejemplo, de los alimentos) y las necesidades urgentes (por ejemplo, el hambre, el deseo sexual y la motivación para el logro) en la reactividad de las señales de CPF. Por ejemplo, en controles sanos jóvenes, el deseo de alimentos imaginados, inducido por una dieta monótona, se asoció con la activación en varias regiones límbicas y paralímbicas, incluida la ACC (área de Brodmann 24) señales de drogas frente a señales neutrales) y para realizar correlaciones de todo el cerebro con los cambios de comportamiento respectivos. Los estudios futuros también podrían comparar la duración y el patrón de activación de CPF después de la exposición aguda al fármaco y después de la exposición a señales condicionadas en los mismos sujetos. Los estudios en individuos no adictos se podrían usar para evaluar el impacto de la privación (por ejemplo, de los alimentos) y las necesidades urgentes (por ejemplo, el hambre, el deseo sexual y la motivación para el logro) en la reactividad de las señales de CPF. Por ejemplo, en controles sanos jóvenes, el deseo de alimentos imaginados, inducido por una dieta monótona, se asoció con la activación en varias regiones límbicas y paralímbicas, incluida la ACC (área de Brodmann 24) Los estudios futuros también podrían comparar la duración y el patrón de activación de CPF después de la exposición aguda al fármaco y después de la exposición a señales condicionadas en los mismos sujetos. Los estudios en individuos no adictos se podrían usar para evaluar el impacto de la privación (por ejemplo, de los alimentos) y las necesidades urgentes (por ejemplo, el hambre, el deseo sexual y la motivación para el logro) en la reactividad de las señales de CPF. Por ejemplo, en controles sanos jóvenes, el deseo de alimentos imaginados, inducido por una dieta monótona, se asoció con la activación en varias regiones límbicas y paralímbicas, incluida la ACC (área de Brodmann 24) Los estudios futuros también podrían comparar la duración y el patrón de activación de CPF después de la exposición aguda al fármaco y después de la exposición a señales condicionadas en los mismos sujetos. Los estudios en individuos no adictos se podrían usar para evaluar el impacto de la privación (por ejemplo, de los alimentos) y las necesidades urgentes (por ejemplo, el hambre, el deseo sexual y la motivación para el logro) en la reactividad de las señales de CPF. Por ejemplo, en controles sanos jóvenes, el deseo de alimentos imaginados, inducido por una dieta monótona, se asoció con la activación en varias regiones límbicas y paralímbicas, incluida la ACC (área de Brodmann 24)58.
Es importante tener en cuenta que, como no hemos revisado la literatura del estriado ventral y, por lo tanto, no se pueden hacer comparaciones directas entre CPF y las respuestas subcorticales a estos estímulos, no podemos inferir, por muy tentador que sea, la actividad de CPF en sí misma puede contribuir a los efectos gratificantes de drogas y estímulos de drogas.
Respuestas a recompensas no relacionadas con drogas
Proponemos que, en individuos con adicción a las drogas, la actividad de CPF en respuesta a recompensas no relacionadas con las drogas es opuesta a los cambios en la actividad de CPF que caracterizan el procesamiento relacionado con las drogas (FIG. 3). Específicamente, en individuos adictos que se encuentran en un estado de deseo, intoxicación, abstinencia o abstinencia temprana, la sensibilidad del CPF a las recompensas no relacionadas con las drogas será notablemente atenuada en comparación con la de los sujetos sanos no adictos. De hecho, la disminución de la sensibilidad a las recompensas no relacionadas con las drogas es un desafío en la rehabilitación terapéutica de pacientes con trastornos por uso de sustancias. Por lo tanto, es importante estudiar cómo responden los individuos adictos a las drogas a los reforzadores no relacionados con las drogas.
Dicha sensibilidad reducida a la recompensa no relacionada con las drogas se ha explicado como una adaptación alostática 59. En esta interpretación, el uso frecuente y de alta dosis de drogas conduce a cambios cerebrales compensatorios que limitan los procesos hedónicos y motivacionales apetitosos («recompensa»), y en su lugar fortalecen los sistemas aversivos (oponentes o «antirrecompensa») 60. Este proceso es similar a la tolerancia, en el cual la sensibilidad a la recompensa disminuye. También es capturado por la hipótesis del proceso de oposición expuesto por Slomon y Corbit 61, 62, que describe la dinámica temporal de las respuestas emocionales opuestas; aquí, el refuerzo negativo (por ejemplo, la abstinencia) prevalece sobre el refuerzo positivo (por ejemplo, alto inducido por drogas) en la transición del uso ocasional de drogas a la adicción. Este proceso es relevante para la reactividad emocional y la regulación de la emoción, que, en la medida en que las emociones se definen como «estados provocados por los refuerzos» 63, se verán afectados por la adicción a las drogas, especialmente durante el procesamiento con tendencia a las drogas, como el ansia y el atracón.
La anhedonia es una característica definitoria de la dependencia de drogas 64, y los criterios para el trastorno depresivo mayor – que incluye anhedonia como un síntoma central – se cumplen por muchos individuos adictos a las drogas (por ejemplo, 50% de los individuos adictos a la cocaína 65). La fuerte asociación entre el estado de ánimo y los trastornos por uso de sustancias no se limita a la depresión 66; por ejemplo, la angustia emocional es un factor de riesgo para la recaída de drogas 67. Sin embargo, la investigación sobre cómo el procesamiento de emociones alteradas está implicado en los trastornos por uso de sustancias se encuentra en su infancia 68, 69, como se explica a continuación (Información complementaria S5 (tabla)).
El dinero es un reforzador abstracto, secundario y generalizable efectivo que adquiere su valor mediante la interacción social, y se usa en el aprendizaje emocional en la experiencia humana cotidiana; el procesamiento comprometido de esta recompensa puede, por lo tanto, apuntar a un mecanismo de aprendizaje emocional socialmente desventajoso en la adicción. Tal déficit, tanto más distintivo dado el fuerte valor motivacional y de excitación que normalmente se asocia con esta recompensa, corroboraría la idea de que, en la adicción, los circuitos de recompensa cerebral son «secuestrados» por las drogas, aunque existe la posibilidad de un déficit preexistente. En el procesamiento de recompensas tampoco se puede descartar.
Un estudio fMRI investigó cómo los individuos y controles adictos a la cocaína respondían a recibir una recompensa monetaria por el desempeño correcto en una atención sostenida y una tarea de elección forzada 70. En los controles, la recompensa monetaria sostenida (ganancia que no varió dentro de los bloques de tareas y que era completamente predecible) se asoció con una tendencia para que la OFC lateral izquierda respondiera de manera gradual (la actividad aumentó monótonamente con la cantidad: ganancia alta> ganancia baja> sin ganancia), mientras que el DLPFC y el ACC rostral respondieron por igual a cualquier monto monetario (ganancia alta o baja> sin ganancia). Este patrón es consistente con el rol de la OFC en el procesamiento de la recompensa relativa, como se documenta en sujetos no humanos 71 y humanos 72 – 76, y con el rol del DLPFC en la atención 77. Los sujetos adictos a la cocaína mostraron señales de fMRI reducidas en la OFC izquierda para una alta ganancia en comparación con los controles y fueron menos sensibles a las diferencias entre las recompensas monetarias en la OFC izquierda y en la DLPFC. Sorprendentemente, más de la mitad de los sujetos adictos a la cocaína calificaron el valor de todas las cantidades monetarias por igual (es decir, US $ 10 = US $ 1000) 78. El ochenta y cinco por ciento de la varianza en estas clasificaciones podría atribuirse a las respuestas laterales a la OFC y al giro frontal medial (y la amígdala) a la recompensa monetaria en los sujetos adictos. Aunque estos hallazgos deben replicarse en un tamaño de muestra más grande y con tareas más sensibles, aun así, sugieren que algunos individuos adictos a la cocaína pueden tener una sensibilidad reducida a las diferencias relativas en el valor de las recompensas. Este «aplanamiento» del gradiente de refuerzo percibido puede ser la base de una sobrevaloración o sesgo hacia las recompensas inmediatas (como una droga disponible) 79 y la depreciación de recompensas mayores pero retrasadas 80, 81, por lo tanto, reduciendo el impulso motivacional sostenido. Estos resultados pueden ser terapéuticamente relevantes, ya que se ha demostrado que el refuerzo monetario en entornos bien supervisados aumenta la abstinencia de las drogas 82, y también puede ser relevante para predecir los resultados clínicos. En línea con esta idea, en una población similar de sujetos, el grado de hipoactivación de dACC en una tarea en la que el desempeño correcto fue remunerado monetariamente correlacionado con la frecuencia de consumo de cocaína, mientras que el grado de hoactivación rostroventral ACC (que se extiende a mOFC) se correlacionó con la tarea supresión del ansia inducida 83. Hubo una asociación inversa de estas ROI de PFC con reactividad de señal en el cerebro medio en sujetos adictos a la cocaína, pero no en sujetos control, lo que implica estas subdivisiones de ACC en la regulación de las respuestas automáticas a las drogas 84.
Cabe señalar que, en los estudios descritos anteriormente, a los sujetos no se les pidió que eligieran entre recompensas monetarias. Predecimos que la elección seguiría de manera similar una función lineal (elección de mayor a menor recompensa) en controles sanos más que en individuos adictos, a quienes esperamos que muestren menos flexibilidad en la elección (elegir la droga en lugar de otros refuerzos), especialmente durante el deseo y el atracón. Los estudios que permiten a los sujetos elegir entre reforzadores se han realizado principalmente en animales de laboratorio. Estos estudios han demostrado que, cuando se les da la opción, los animales previamente expuestos a las drogas eligen la droga en lugar de la novedad 85, el comportamiento materno adecuado 86 e incluso la comida 87 – 89, lo que indica que la exposición al medicamento puede disminuir el valor percibido de las recompensas naturales, incluso aquellas que son necesarias para la supervivencia. En un estudio reciente de neuroimagen humana en el que los sujetos podían ganar cigarrillos o dinero, los fumadores ocasionales estaban más motivados para obtener dinero que los cigarrillos, mientras que los fumadores dependientes hicieron esfuerzos similares para ganar dinero o cigarrillos 90. Se observó una interacción similar de grupo por recompensa en la OFC derecha, DLPFC bilateral y CAC izquierda, de manera que en los fumadores ocasionales estas regiones mostraron una mayor actividad a los estímulos que predicen una recompensa monetaria creciente que a los estímulos que predicen una recompensa de cigarrillos, mientras que los fumadores dependientes mostraron No hay diferencias significativas en dicha actividad cerebral anticipatoria. Estas regiones también mostraron una mayor activación del dinero en el caso ocasional que en los fumadores dependientes 90.
Estos resultados, junto con los resultados de comportamiento en las pruebas neuropsicológicas en individuos adictos a la cocaína 91 , 92 (ver también RECUADRO 2), contribuyen a nuestra comprensión de cómo las preferencias de recompensa relativas pueden cambiar en la adicción, de modo que la preferencia por la droga compite con (y algunas veces supera) preferencia por otros reforzadores, con una disminución concomitante en la capacidad de asignar valores relativos a recompensas no relacionadas con las drogas.
Reactividad emocional
Varios estudios que se revisaron anteriormente compararon las respuestas de PFC a estímulos no específicos, pero que provocaban emociones, con respuestas a indicaciones relacionadas con la preocupación (por ejemplo, relacionadas con las drogas) 25, 26, 28, 46, 47 (Información complementaria S3 (tabla)). El PFC fue hiperactivo en respuesta a las imágenes de todas las categorías emocionales en sujetos adictos al alcohol 28, el PFC anterior fue hipoactivo en respuesta a imágenes agradables en individuos con adicción a la heroína 26, y en pacientes con trastornos de la alimentación. Las respuestas del PFC a las imágenes aversivas fueron normales 46, 47. Así, en contraste con las predicciones de nuestro modelo (HIGO. 3), en ninguno de estos estudios hubo diferencias en la respuesta de PFC entre señales relacionadas con el fármaco y afectivas, pero no relacionadas con el fármaco. Este resultado, y la variabilidad en el patrón de resultados, podrían atribuirse, entre otros factores, al pequeño número de estudios, las diferencias entre los estudios (como el tamaño de las muestras, la droga primaria de abuso y la duración de la abstinencia) y la sensibilidad de los pacientes. Medidas utilizadas. Los estudios futuros se beneficiarían con el uso de grabaciones potenciales relacionadas con eventos o electroencefalografía, que tienen una resolución temporal mucho mayor que la RMf o la PET.
Surge una imagen más clara cuando los estudios incorporan el procesamiento emocional en las tareas cognitivo-conductuales (Información complementaria S5 (tabla)). Por ejemplo, cuando se requiere empatizar con un protagonista en una serie de caricaturas, cada una con una historia corta, los individuos adictos a la metanfetamina proporcionaron menos respuestas correctas que los controles a la pregunta «¿qué hará que el personaje principal se sienta mejor?» 93. En comparación con los sujetos de control, los individuos adictos también mostraron hipoactivación en OFC (e hiperactivación en DLPFC) al responder esta pregunta. Con la excepción de un estudio en individuos abstinentes adictos a la heroína 94 Otros estudios similares también informaron diferencias entre los grupos adictos y de control en las respuestas de PFC a tareas que requieren el procesamiento de estímulos emocionales, como rostros, palabras o escenas complejas. Por ejemplo, cuando los hombres con adicción al alcohol juzgaron la intensidad de cinco expresiones faciales, las expresiones negativas se asociaron con activaciones más bajas en el ACC izquierdo, pero activaciones más altas en el DLPFC izquierdo y dACC derecho en comparación con los controles 95. Además, en comparación con los controles sanos, los usuarios de cocaína mostraron Hipoactivaciones de PFC dorsomedial y ACC mientras realizaban una tarea de discriminación de letras durante la presentación de un conjunto de imágenes agradables (versus neutrales) e hiperactivaciones en la DLPFC bilateral durante la presentación de desagradables (versus agradables) fotos 96. De manera similar, en comparación con los controles sanos, los fumadores de marihuana mostraron hipoactivaciones de la CAC izquierda, e hiperactivaciones del DLPFC derecho y del giro frontal inferior en respuesta a la presentación de caras enojadas enmascaradas (frente a caras neutrales); las respuestas correctas de ACC se correlacionaron positivamente con la frecuencia de consumo de drogas y las respuestas bilaterales de ACC se correlacionaron con los niveles de cannabinoides en la orina y el consumo de alcohol 97. Por el contrario, el dACC izquierdo fue hiperactivo en sujetos dependientes de la metanfetamina en comparación con los controles al juzgar la expresión emocional en las caras en una tarea de emparejamiento de afecto (versus la forma de figuras abstractas) y esto se asoció con más hostilidad auto informada y sensibilidad interpersonal en Los sujetos adictos 98.
En conjunto, estos estudios indican que el DLPFC es en su mayoría hiperactivo durante el procesamiento de la emoción en individuos adictos en comparación con los sujetos de control, especialmente para las emociones negativas. El ACC muestra resultados mixtos, aunque con más estudios que muestran hipoactividad que hiperactividad. Es posible que la hiperactividad de la DLPFC compense la hipoactividad del ACC, lo que explicaría la falta de diferencia en el desempeño de la tarea entre los drogadictos y los controles sanos en la mayoría de estos estudios. Se pueden observar conductas desventajosas y / o impulsivas durante los desafíos de mayor excitación emocional, como el estrés, el deseo o tareas más difíciles. Claramente, los roles de estas regiones en relación con el modelo propuesto (FIG. 3) necesitan ser mejor entendidos. Es posible que, al reclutar prematuramente la función ejecutiva de PFC de orden superior (mediada por el DLPFC), la activación emocional negativa aumente el riesgo de consumo de drogas en personas adictas, particularmente en situaciones que ejercen una presión adicional sobre los recursos limitados de control cognitivo. Esta interpretación es consistente con la competencia entre los procesos relacionados con medicamentos y no relacionados con los medicamentos y entre los procesos «en frío» y «en caliente» en el modelo (Figura 3c). Aunque varios de los estudios anteriores utilizaron estímulos con valencia negativa, una pregunta persistente es si la sensibilidad alterada a los reforzadores no farmacológicos en individuos adictos también se aplica a los reforzadores negativos como la pérdida de dinero. Los estudios en animales muestran que los sujetos «adictos» manifiestan una búsqueda persistente de drogas, incluso si la droga está asociada con una descarga eléctrica 99. En humanos, se ha informado 100 hipoactivación en el PFC ventrolateral derecho en fumadores durante la pérdida monetaria y en jugadores durante la ganancia monetaria (Información complementaria S5 (tabla)). Aunque es evidente que se necesitan más estudios, la implicación de una menor sensibilidad a los reforzadores negativos en la adicción tiene implicaciones prácticas, ya que, además de los reforzadores positivos (como los vales y privilegios), los reforzadores negativos (como el encarcelamiento) se utilizan cada vez más en el manejo de drogadictos. Las intervenciones podrían optimizarse seleccionando el tipo y la dosis más efectivos de refuerzo. Los estudios futuros también podrían ayudar a determinar si las personas adictas pueden recurrir a tomar drogas porque se aburren, se sienten frustradas, enojadas o temerosas fácilmente, tal vez como resultado de un funcionamiento alterado del PFC. Umbral bajo para experimentar cualquiera de estas emociones, o la incapacidad de mantener un comportamiento dirigido a un objetivo (por ejemplo, completar una tarea aburrida) al experimentar estas emociones, se puede asociar con un control inhibitorio deteriorado (es decir, impulsividad mejorada) como se analiza a continuación. En los individuos adictos a la cocaína, la actividad de PFC se habitúa prematuramente a la presentación repetida de una tarea de atención sostenida de incentivo 101, que podría ser una medida de la sostenibilidad comprometida del esfuerzo y resultar en una participación inadecuada en las actividades de tratamiento.
Control inhibitorio en la adicción.
La adicción a las drogas se caracteriza por alteraciones leves, sin embargo, penetrantes, cognitivas 102 que pueden acelerar su curso, amenazar la abstinencia sostenida 103 o aumentar el desgaste del tratamiento 104, 105. El CPF es esencial para muchos de estos procesos cognitivos, incluida la atención, la memoria de trabajo, la toma de decisiones y el descuento por demora (TABLA 1), todos los cuales están comprometidos en individuos adictos, según se revisa en otra parte 106. Otra función cognitiva importante del PFC es el autocontrol, y aquí nos centramos en el papel del PFC en este proceso en la adicción (Información complementaria S6 (tabla)). El autocontrol se refiere, entre otras operativas, a la capacidad de una persona para guiar o detener un comportamiento, particularmente cuando el comportamiento puede no ser óptimo o ventajoso, o si se percibe como algo incorrecto. Esto es pertinente para la adicción, ya que, a pesar de cierta conciencia de las consecuencias devastadoras de las drogas (consulte también la sección a continuación sobre la conciencia de la enfermedad en la adicción), los individuos adictos a las drogas muestran una capacidad deficiente para inhibir el consumo excesivo de drogas. El control inhibitorio deteriorado, que es una operación clave en el autocontrol, también es probable que contribuya a la participación en actividades delictivas para obtener el medicamento y para sustentar la regulación deficiente de las emociones negativas, como se sugirió anteriormente. Estas deficiencias también podrían predisponer a los individuos a la adicción. De acuerdo con informes anteriores 107, el autocontrol de los niños durante su primera década de vida predice la dependencia de sustancias en su tercera década de vida 108.
Tareas de ir / no ir y detener el tiempo de reacción de la señal
Las tareas que se usan a menudo para medir el control inhibitorio son la tarea de ir / no ir y la tarea de tiempo de reacción de la señal de parada (SSRT). En la tarea de ir / no ir, los individuos adictos a la cocaína mostraron más errores de omisión y comisión que los controles y esto se ha atribuido a la hipoactivación en dACC durante los ensayos de detención 109. En otro estudio, este déficit de comportamiento inhibitorio en los usuarios de cocaína se vio agravado por una mayor carga de memoria de trabajo; de nuevo, la hipoactivación de dACC se asoció con un desempeño de tareas deficiente 110. De manera similar, los hombres adictos a la heroína mostraron tiempos de reacción más lentos en la tarea de ir / no ir, junto con la hipoactivación en el ACC y el PFC medial 111. Los resultados de la SSRT son más difíciles de interpretar. Por ejemplo, el ACC fue hipoactivo durante las inhibiciones de respuesta exitosas en comparación con las inhibiciones de respuesta fallidas en hombres adictos a la cocaína, y su comportamiento de comportamiento fue similar al de los controles 112. El ACC también fue hipoactivo durante el ajuste cuidadoso del comportamiento y el riesgo de asumir esta tarea en alcohólicos abstinentes, particularmente en sujetos con mayor necesidad de alcohol en el momento de la exploración por resonancia magnética funcional (IRMR) 113. Por el contrario, el ACC fue hiperactivo durante los errores de inhibición 113, posiblemente porque los alcohólicos abstinentes ejercieron una mayor atención en el monitoreo de la señal de parada que en los controles, una función que está asociada con el ACC. El aumento de la actividad en otras regiones del PFC también se informó en fumadores de cigarrillos después de una abstinencia de 24 horas, pero (en contraste con la expectativa de un aumento de la activación regional) la precisión se redujo en 114 (Información complementaria S4 (tabla)). La gran variabilidad en los resultados de estos estudios es posiblemente causada por las diferencias en los análisis, el tipo de comparación y las diferencias de rendimiento entre los grupos, además de otras variables. Sin embargo, surge un patrón en el que el dACC es hipoactivo durante estas tareas de control inhibitorio, y esta hipoactividad se asocia principalmente con un desempeño deficiente, particularmente con duraciones de abstinencia más cortas. Intervenciones cognitivo-conductuales dirigidas pueden aliviar esta disfunción. Por ejemplo, la información informativa (como proporcionar una advertencia de un ensayo inminente de no ir) mejoró el control inhibitorio en una tarea de ir / no ir, y esto se correlacionó con una mayor activación de ACC en individuos adictos a la metanfetamina 115. Tales intervenciones cognitivas / conductuales podrían usarse como ejercicios de rehabilitación neuronal y combinarse con la administración simultánea de medicamentos, como se explica a continuación.
Tareas de Stroop
El control inhibitorio también puede evaluarse mediante la palabra de color tarea Stroop 116. Un rendimiento más lento y más errores durante las pruebas incongruentes en esta tarea son un sello distintivo de la disfunción de PFC. La investigación en neuroimagen ha demostrado que el dACC y el DLPFC están involucrados en esta tarea 117 – 119, con roles distintos para estas regiones en la detección de conflictos (dACC) y la resolución (DLPFC) 120.
Los estudios que utilizan la tarea de color-palabra Stroop en individuos adictos reportan resultados que en su mayoría reflejan los reportados anteriormente. Por ejemplo, los abusadores de cocaína tenían un FCB más bajo en el dACC izquierdo y el DLPFC derecho durante los ensayos incongruentes en comparación con los ensayos congruentes, mientras que el ACC derecho mostró el patrón opuesto; además, la activación derecha de ACC se correlacionó negativamente con el consumo de cocaína 121 (Información complementaria S6 (tabla)). En los hombres que consumían marihuana, se notificó una FBC más baja durante esta tarea en varias regiones de CPF, incluido el ACC perigenual, el PFC ventromedial y el DLPFC 122. Los sujetos dependientes de la metanfetamina también mostraron hipoactivaciones en la red de control inhibitorio, incluyendo dACC y DLPFC mientras realizaban esta tarea 123. Consistente con el impacto de la abstinencia en la tarea de ir / no ir reportada por encima de 114, los fumadores de cigarrillos que se probaron después de una abstinencia de 12 horas redujeron los tiempos de reacción, aumentaron el dACC y redujeron las respuestas de la DLPFC a los ensayos incongruentes en el color. word Stroop task 124 (Información complementaria S4 (tabla)). Es importante destacar que un estudio de resonancia magnética nuclear demostró que la activación del PFC ventromedial (Brodmann áreas 10 y 32) durante una tarea de Stroop de color-palabra realizada 8 semanas antes del inicio del tratamiento predijo el resultado del tratamiento en individuos adictos a la cocaína 125.
En la variante emocional de esta tarea, las palabras de color se sustituyen por palabras o imágenes emocionales que se relacionan con el área de preocupación de un individuo en particular, como las palabras relacionadas con el alcohol para personas adictas al alcohol. Aunque tanto las pruebas Stroop clásicas como las emocionales implican la necesidad de suprimir las respuestas a la información de estímulo que distrae mientras se mantiene selectivamente la atención sobre la propiedad del estímulo que se necesita para completar la tarea, solo la tarea de Stroop emocional utiliza la relevancia emocional como distractor. Tales diseños de Stroop emocionales pueden potencialmente demarcar aún más la actividad de CPF alterada en la adicción: ¿es generalizable a cualquier tipo de conflicto u ocurre específicamente durante conflictos en un contexto relacionado con las drogas?
Un estudio de resonancia magnética funcional en usuarios de estimulantes mostró un sesgo de atención a las palabras relacionadas con las drogas: los individuos adictos, pero no los controles, mostraron más sesgos de atención a las palabras relacionadas con las drogas (medida como la latencia de respuesta media de los colores correctamente identificados de las palabras relacionadas con las drogas menos la mediana La latencia de respuesta de los colores correctamente identificados de palabras neutras coincidentes), que se correlacionó con las respuestas mejoradas de PFC ventral izquierdo. Dichas respuestas no se observaron para la tarea de color-palabra Stroop 126. De manera similar, las imágenes relacionadas con las drogas amplificaron las respuestas de dACC a la información relevante para la tarea en fumadores de cigarrillos 127. Estos hallazgos sugieren que, en la adicción, se necesitan más recursos de arriba hacia abajo para centrarse en las tareas cognitivas cuando las señales relacionadas con las drogas están presentes como distractores (por lo tanto, desvían la atención) durante la tarea. Conflicto con estos y otros resultados 128 son estudios en usuarios actuales de cocaína, en los cuales las palabras relacionadas con las drogas no se asociaron con un rendimiento más lento o con más errores 83, 129. Esta disparidad podría estar relacionada con el diseño de la tarea o el estado de búsqueda de tratamiento de los participantes del estudio; predecimos que el mayor conflicto entre las palabras relacionadas con las drogas y las palabras neutrales caracteriza a las personas que intentan abstenerse de consumir drogas. La evidencia de tal efecto en los fumadores de cigarrillos se publicó recientemente 130.
Efectos de la administración de drogas durante las tareas de control inhibitorio.
Las deficiencias en la regulación emocional y el control inhibitorio en individuos adictos y el aumento de la actividad de PFC mediante la administración directa de medicamentos (ver más arriba y la información complementaria S2 (tabla)) en conjunto podrían apoyar la hipótesis de automedicación 131, 132. De acuerdo con esta hipótesis, la autoadministración de medicamentos, y los aumentos asociados en la actividad de PFC, mejoran los déficits emocionales y cognitivos que están presentes en las personas adictas a los medicamentos. Tal efecto de automedicación ha sido reconocido previamente por la comunidad de tratamiento, como lo demuestra el uso de la metadona (un opioide sintético) como una terapia estándar de sustitución con agonistas para la dependencia de la heroína. En un estudio de resonancia magnética nuclear, observar señales relacionadas con la heroína se asoció con menos antojo durante una dosis posterior a la de una sesión de metadona previa a la dosis en individuos adictos a la heroína, con disminuciones concomitantes en las respuestas relacionadas en el OFC 133 bilateral (Información complementaria S4 (tabla)). El apoyo empírico está comenzando a acumularse por un efecto similar en individuos adictos a la cocaína. Por ejemplo, la cocaína intravenosa (que aumenta los niveles de dopamina extracelular) en los usuarios de cocaína mejoró el control inhibitorio en una tarea de ir / no ir, y esto se asoció con la normalización de la actividad de ACC y la activación mejorada del DLPFC derecho durante la tarea 134. La MPH intravenosa (que también aumenta los niveles de dopamina extracelular) mejoró de manera similar el rendimiento en la prueba SSRT en los consumidores de cocaína, y esto se correlacionó positivamente con la activación relacionada con la inhibición de la corteza frontal media izquierda y se correlacionó negativamente con la actividad en el CPF ventromedial; después de la MPH, la actividad en ambas regiones mostró una tendencia a la normalización 135. Un estudio de PET mostró que la MPH oral atenuaba el metabolismo reducido en las regiones del cerebro límbico, incluyendo la OFC lateral y la DLPFC, que seguían la exposición a señales relacionadas con la cocaína en individuos con adicción a la cocaína 136. También disminuyó los errores de comisión, una medida común de impulsividad, durante una tarea de Stroop emocional relevante para las drogas, tanto en individuos adictos a la cocaína y en controles, como en los individuos adictos, esta disminución se asoció con la normalización de la activación en el CAC rostroventral (extensión al mOFC) y dACC; La activación relacionada con la tarea de dACC antes de la administración de MPH se correlacionó con el uso más corto de alcohol de por vida 137 ( FIG. 4). Si bien queda por estudiar si los efectos noradrenérgicos de la MPH contribuyen a sus efectos de «normalización» en los consumidores de cocaína, estos resultados sugieren que los efectos del MPH que aumentan la dopamina podrían usarse para facilitar cambios en el comportamiento de los individuos adictos (por ejemplo, mejorar el autocontrol), particularmente si el tratamiento de MPH se combina con intervenciones cognitivas específicas.
Cabe señalar que el efecto de los agonistas de la dopamina en la normalización de las respuestas del comportamiento del cerebro a los desafíos emocionales o de control cognitivo puede depender de los patrones de uso compulsivo de drogas 126 u otras diferencias individuales, como el autocontrol de referencia y el uso de drogas de por vida, pero Las posibilidades quedan por estudiar en muestras de mayor tamaño. Además, las sondas no dopaminérgicas (por ejemplo, agonistas colinérgicos o receptores de AMPA) pueden ofrecer objetivos farmacológicos adicionales para el tratamiento 138 de la adicción a la cocaína.
En resumen, los resultados de los estudios sobre el control inhibitorio en la adicción a las drogas sugieren que existe una hipoactividad dACC y un control inhibitorio deficiente en los individuos adictos a las drogas. La actividad de PFC mejorada se ha informado después de la abstinencia a corto plazo, después de la exposición a señales relacionadas con el fármaco y al propio fármaco (o agentes farmacológicos similares). Sin embargo, aunque la exposición a drogas también se asocia con un mejor desempeño en estas tareas cognitivas, la abstinencia a corto plazo y la exposición a señales relacionadas con las drogas tienen el resultado opuesto en el desempeño de tareas. Visto en el contexto del modelo propuesto (FIG. 3), aunque las drogas de abuso ofrecen un alivio temporal, la automedicación crónica con estas drogas tiene consecuencias a largo plazo (mecanismos reducidos de control inhibitorio y trastornos emocionales asociados) que no pueden aliviarse con la abstinencia a corto plazo y que son propensos a ser reavivados tras la exposición a señales relacionadas con las drogas. La normalización de estas funciones, utilizando intervenciones farmacológicas y cognitivo-conductuales de base empírica y dirigidas, en combinación con los refuerzos relevantes, debe convertirse en un objetivo en el tratamiento de la adicción.
La conciencia de la enfermedad en la adicción
La capacidad de comprensión de nuestro mundo interno (que abarca la interocepción pero que se extiende a una autoconciencia emocional, motivacional y cognitiva de orden superior) depende en parte del CPF. Dados los impedimentos en la función de CPF en personas con adicción revisada anteriormente, es posible que un conocimiento restringido de la magnitud del deterioro conductual o de la necesidad de tratamiento pueda ser la base de lo que tradicionalmente se ha atribuido a la «negación» en la adicción a las drogas, es decir, la suposición de que el paciente adicto es capaz de comprender completamente sus déficits, pero decide ignorarlos puede ser errónea. De hecho, los estudios han sugerido recientemente que las personas adictas no son plenamente conscientes de la gravedad de su enfermedad (es decir, su comportamiento de búsqueda y consumo de drogas y sus consecuencias) y esto puede estar asociado con déficits en la red de control139.
Varios estudios han proporcionado evidencia de una disociación entre la autopercepción y el comportamiento real en la adicción. Por ejemplo, en controles saludables, la velocidad y la precisión de las respuestas para una condición monetaria alta en comparación con una señal neutral en una tarea de atención sostenida por elección forzada y remunerada monetariamente se correlacionó con el compromiso autoinformado en la tarea; por el contrario, los informes de los sujetos de la cocaína sobre el compromiso de la tarea se desconectaron de su desempeño real de la tarea, lo que indica discordancia entre la motivación autoinformada y el comportamiento orientado hacia los objetivos 70 . Uso de una tarea desarrollada recientemente en la que los participantes seleccionaron sus imágenes preferidas de cuatro tipos de imágenes y luego informaron lo que pensaron que era su tipo de imagen más seleccionada 91, la discordancia entre el autoinforme y la elección real, lo que indica una percepción deficiente del propio comportamiento de elección, fue más grave en los consumidores actuales de cocaína, aunque también fue perceptible en los usuarios abstinentes, en los cuales se correlacionó con la frecuencia del consumo reciente de cocaína 92 .
Un mecanismo subyacente de esta disociación puede ser un desacoplamiento de las respuestas de comportamiento y autonómicas durante el aprendizaje de inversión, como se ha demostrado que ocurre después de la lesión por OFC en monos 140. Existe cierta evidencia de disociaciones neural-conductuales similares también en los humanos. En un estudio de potencial relacionado con un evento que utiliza la tarea informada arriba.70 Los sujetos de control mostraron respuestas electrocorticales alteradas y tiempos de reacción en la condición de alto dinero en comparación con la condición de señal neutral, y estas dos medidas de atención motivada se interrelacionaron. Este patrón no se observó en el grupo adicto a la cocaína, en el que la capacidad de responder con precisión al dinero (es decir, cuanto mayor es la flexibilidad conductual de este reforzador), se correlacionó negativamente con la frecuencia del consumo reciente de cocaína 141. Otro estudio mostró que, en una tarea de juego, las elecciones de los sujetos de control se guiaban por errores reales y ficticios, mientras que los fumadores de cigarrillos solo se guiaban por los errores reales que habían cometido, aunque los errores ficticios indujeron respuestas neuronales robustas 142, nuevamente señalando disociaciones neural-conductuales en la adicción. En el modelo propuesto (Figura 3), este mecanismo está representado por una entrada disminuida de las regiones de control cognitivo de orden superior a las regiones que están asociadas con el procesamiento emocional y las respuestas condicionadas. Es importante destacar que, en los seres humanos, esta disociación neural-conductual se puede validar comparando los autoinformes de los pacientes con los de los informantes 137, como los miembros de la familia o los proveedores de tratamiento, o con medidas objetivas de rendimiento en las pruebas neuropsicológicas 143. Es importante recordar que las medidas de autoinforme brindan una visión importante de tales disociaciones, pero dadas las limitaciones de los autoinformes, el desarrollo de medidas más objetivas de conocimiento y conciencia es crucial tanto para la investigación como para fines clínicos. Dos medidas prometedoras son la concientización de errores y afectan la correspondencia Se encontró que la concientización de errores en una tarea de ir / no ir se redujo en los jóvenes consumidores de marihuana y esto se asoció con reducciones en el DLPFC bilateral y el CAC correcto, y con un mayor uso actual de drogas 144. En sujetos dependientes de la metanofetamina, el PFC ventrolateral bilateral fue hipoactivo durante el emparejamiento del afecto y esto se asoció con más alexitimia autoinformada 145. Como un mejor conocimiento de la gravedad del uso de drogas predijo la abstinencia real hasta 1 año después del tratamiento en alcohólicos 146, esta línea de investigación en ciernes podría mejorar enormemente nuestra comprensión de la recaída en la adicción a las drogas, mejorando potencialmente los enfoques de intervención actualmente disponibles, por ejemplo, mediante dirigidos a individuos adictos que han reducido la autoconciencia para intervenciones a medida.
Estudio de limitaciones y direcciones futuras.
La principal limitación de esta Revisión es nuestro enfoque selectivo en el PFC a costa de excluir todas las demás regiones cerebrales corticales y estructuras subcorticales. La arquitectura de apoyo a la función ejecutiva de orden superior y el control de arriba hacia abajo es compleja y se cree que implica varias redes funcionales que incluyen, además de la PFC, otras regiones como la corteza parietal superior, insula, el tálamo y el cerebelo 147. En consecuencia, y también dadas las limitaciones inherentes de los estudios transversales de neuroimagen humana, debe evitarse la atribución de causalidad, es decir, el PFC puede no conducir directamente a los déficits descritos en esta revisión. Los futuros metaanálisis en los que se explora la interrupción de estas redes funcionales en la adicción deben estar imbuidos de los resultados de estudios mecanizados en animales de laboratorio.
Un problema notable con muchos de los estudios revisados se refiere a su uso de los análisis de ROI funcionales que a veces carecen de las correcciones estadísticas más estrictas de los análisis de todo el cerebro. Por ejemplo, para superar los problemas de baja potencia, los resultados informados a veces se restringen a análisis post-hoc en regiones que mostraron resultados significativos en todas las materias para todas las condiciones de la tarea; No se realizan de manera consistente los análisis de los efectos principales (por ejemplo, grupo o tipo de estímulo) o de interacción, o de las correlaciones con el desempeño de la tarea o los puntos finales clínicos, de forma integral. Por lo tanto, tales resultados de ROI podrían representar un error de Tipo I, pero también podrían pasar por alto los sustratos neuronales clave que están involucrados en el fenómeno bajo investigación, por ejemplo, el deseo o el control del deseo. ROI anatómicas definidas a priori 148, 149, lo que también podría ayudar a estandarizar la nomenclatura de las ROI entre los estudios. Otros problemas comunes se relacionan con la presentación incompleta de los datos reales (como no proporcionar la media y la varianza, o no proporcionar diagramas de dispersión cuando se reportan correlaciones), lo que puede oscurecer la dirección de un efecto (activación versus desactivación), lo que potencialmente aumenta la variabilidad en Resultados publicados (por ejemplo, una hiperactivación podría referirse a activaciones más altas o desactivaciones más bajas desde la línea de base). En resumen, este campo se beneficiaría de la estandarización, de los procedimientos relacionados con imágenes, tareas, análisis y caracterización de sujetos, que facilitaría la interrelación de los hallazgos. La estandarización también es crucial para permitir la integración de conjuntos de datos de diversos laboratorios; dicha combinación de datos será particularmente importante para los estudios genéticos que tienen como objetivo comprender la interacción entre los genes, el desarrollo cerebral, la función cerebral y los efectos de los medicamentos en estos procesos. Por ejemplo, la creación de grandes conjuntos de datos de imágenes será importante para comprender cómo los genes asociados con la vulnerabilidad a la adicción afectan al cerebro humano después de exposiciones agudas y repetidas a drogas. Además, la capacidad de integrar grandes conjuntos de datos de imágenes, como se ha hecho recientemente para las imágenes de resonancia magnética de la conectividad funcional en reposo la creación de grandes conjuntos de datos de imágenes será importante para comprender cómo los genes asociados con la vulnerabilidad a la adicción afectan al cerebro humano después de exposiciones agudas y repetidas a medicamentos. Además, la capacidad de integrar grandes conjuntos de datos de imágenes, como se ha hecho recientemente para las imágenes de resonancia magnética de la conectividad funcional en reposo150 – permitirá una mejor comprensión de la neurobiología de la adicción que en el futuro puede servir como biomarcador para guiar el tratamiento.
Aunque hay algunas excepciones (que implican el PFC correcto, particularmente el ACC y el DLPFC, en los procesos inhibitorios compensatorios), los datos revisados aquí no muestran un patrón claro que indique la lateralización de los cambios cerebrales en individuos adictos. Sin embargo, la lateralización no fue el foco de investigación en ninguno de los estudios revisados. Dado que existe evidencia de una lateralidad interrumpida durante el uso de los dedos en los consumidores de cocaína 151, se necesitan estudios que investiguen específicamente la lateralización de PFC en iRISA en adicción. Además, existen claras diferencias de género en las respuestas a las drogas y en la transición a la adicción, y los estudios de imágenes están aumentando nuestra comprensión de las características sexualmente dimorfas del cerebro humano. Sin embargo, hasta el momento, pocos estudios bien controlados se han centrado en las diferencias de sexo en el papel de la CPF en la adicción; en cambio, muchos estudios utilizan sujetos femeninos o masculinos (en su mayoría varones). También se necesitan estudios para explorar los efectos potencialmente moduladores de otras características individuales; de particular interés son el impacto de los trastornos comórbidos (por ejemplo, la depresión puede exacerbar los déficits en individuos adictos152) y la frecuencia del uso de drogas y la duración de la abstinencia (por ejemplo, la cocaína puede reducir o enmascarar las deficiencias cognitivas 153 o emocionales 154 subyacentes de la cocaína en individuos adictos. Los estudios longitudinales permitirían el examen de estos temas, que son de particular importancia para quienes se abstienen de consumir drogas con la esperanza de que el funcionamiento del PFC se recupere. Además, la comparación entre los diferentes tipos de sustancias de abuso permitiría la diferenciación entre los factores que son específicos de ciertas drogas y los factores que podrían ser comunes en las poblaciones de adicciones. En lugar de tratar la heterogeneidad de los cambios neuronales y de comportamiento en la adicción como ruido, los estudios podrían explorarlo con el objetivo de responder preguntas clave: ¿es la disfunción de PFC en iRISA más prominente en ciertos individuos adictos que en otros? ¿La automedicación impulsa el consumo de drogas en algunas personas más que en otras? ¿De qué manera el uso de drogas concomitantes, que es más la regla que la excepción (por ejemplo, la mayoría de los alcohólicos son adictos a la nicotina), afectan la neurobiología en la adicción? ¿Cuál es la implicación de esta variabilidad en el resultado del tratamiento y la recuperación? Más importante aún, ¿cómo podemos usar estos resultados de laboratorio en el funcionamiento de PFC en adicción para informar el diseño de intervenciones de tratamiento efectivas?
Resumen y conclusiones
En general, los estudios de neuroimagen han revelado un patrón emergente de disfunción generalizada de CPF en individuos adictos a las drogas que se asocia con resultados más negativos: más consumo de drogas, peor desempeño en las tareas relacionadas con CPF y mayor probabilidad de recaída. En individuos adictos a las drogas, la activación generalizada de CPF al tomar cocaína u otras drogas y al presentar señales relacionadas con las drogas se reemplaza por la hipoactividad generalizada de CPF durante la exposición a desafíos cognitivos y emocionales de orden superior y / o durante la abstinencia prolongada cuando no se estimula. Los roles de CPF que son más pertinentes para la adicción incluyen el autocontrol (es decir, la regulación emocional y el control inhibitorio) para terminar acciones que no son ventajosas para el individuo, la atribución de prominencia y el mantenimiento de la excitación motivacional que es necesario para involucrarse en conductas orientadas a los objetivos y la autoconciencia. Aunque la actividad entre las regiones de CPF es altamente integrada y flexible, por lo que cualquier región está involucrada en múltiples funciones, el CPF dorsal (incluido el dACC, el DLPFC y el giro frontal inferior) se ha implicado predominantemente en el control de arriba hacia abajo y las funciones metacognitivas. , el CPF ventromedial (incluido el ACC subgenual y el mOFC) en la regulación de las emociones (incluido el condicionamiento y la asignación de atención de incentivo a las drogas y los estímulos relacionados con la droga), y el CPF ventrolateral y la OFC lateral en las tendencias de respuesta automática e impulsividad (TABLA 1 ). La disfunción de estas regiones CPF puede contribuir al desarrollo del deseo, el uso compulsivo y la «negación» de la enfermedad y la necesidad de tratamiento, síntomas característicos de la adicción a las drogas. Esta disfunción de CPF puede en algunos casos preceder al uso de drogas y conferir vulnerabilidad para desarrollar trastornos por uso de sustancias (CUADRO 3). Independientemente de la dirección de la causalidad, los resultados de los estudios de neuroimagen que se revisan aquí sugieren la posibilidad de que biomarcadores específicos puedan ser seleccionados para fines de intervención. Por ejemplo, tal vez estas anomalías de CPF podrían usarse para identificar a los niños y adolescentes que se beneficiarían más de los esfuerzos intensivos de prevención del abuso de drogas, y tal vez los medicamentos pueden mejorar estos déficits y ayudar a las personas adictas a participar en un tratamiento de rehabilitación.
Cuadro 3 Vulnerabilidad y predisposición al uso de drogas
Los estudios sobre cómo las vulnerabilidades premórbidas, como la exposición prenatal a medicamentos, los antecedentes familiares o los polimorfismos genéticos seleccionados y sus interacciones, afectan la función de la corteza prefrontal (CPF) son cruciales para el diseño de futuras intervenciones y posiblemente los esfuerzos de prevención; estos estudios resaltan la importancia de identificar biomarcadores claros de vulnerabilidad al uso de drogas y la adicción. Por ejemplo, la reducción del flujo sanguíneo cerebral global absoluto (CBF) (–10%) y la mejora del CBF relativo en el CPF dorsolateral (DLPFC) (9%) y la corteza cingulada anterior (ACC) (12%) se informó en adolescentes con exposición prenatal a la cocaína 201. También se informó un PFC hiperactivo en usuarios jóvenes de MDMA 202, marihuana 203 o alcohol 204 durante la tarea Ir / No ir, en la que se realizaron normalmente (Información complementaria S6 (tabla)). De manera similar, en comparación con los niños control que tenían padres alcohólicos pero eran resistentes, los niños que tenían padres alcohólicos y eran vulnerables al consumo de alcohol (clasificado según el nivel de consumo de alcohol durante el transcurso de la adolescencia) tenían un PFC dorsomedial derecho hiperactivo, mientras que la corteza orbitofrontal bilateral (OFC) era hipoactiva, a pesar de la falta de diferencias de comportamiento cuando se leen palabras emocionales en silencio. En toda la muestra, dicha hiperactividad de PFC dorsomedial se asoció con síntomas más externos y con agresión 205 (Información complementaria S5 (tabla)). Por lo tanto, dichos cambios en la actividad de PFC pueden ser compensatorios en el corto plazo (como lo demuestra el desempeño de tareas iguales), pero en el largo plazo pueden promover el abuso de sustancias y la adicción en estos individuos, aunque esto aún no se ha determinado.
El mecanismo que subyace a dicha vulnerabilidad, o que confiere protección contra el desarrollo de una adicción, puede implicar una neurotransmisión dopaminérgica alterada. Por ejemplo, la disponibilidad del receptor D2 de la dopamina estriatal y el metabolismo regional de la PFC fueron mayores en los miembros jóvenes y no afectados de familias alcohólicas que en los sujetos sin tal historial familiar, que es lo contrario a los resultados comúnmente informados en individuos adictos (RECUADRO 2; ver Información complementaria S7 (tabla)) 206. Los individuos con antecedentes familiares de abuso de alcohol informaron una emocionalidad positiva más baja, y esto se asoció con una menor disponibilidad del receptor D2 de la dopamina del estriado y un menor metabolismo de la OFC. Por lo tanto, es posible que la mayor disponibilidad del receptor D2 de dopamina y la actividad metabólica mejorada en PFC en individuos con antecedentes familiares de abuso de alcohol aumenten el nivel de emocionalidad positiva, aunque esto no obstante se mantuvo por debajo del nivel en controles sanos, a niveles que pueden tener protegió a estos individuos contra el desarrollo de la adicción. También es posible que se necesiten condiciones óptimas para el mantenimiento de dicha protección, y que las condiciones subóptimas (por ejemplo, el estrés crónico) puedan exponer a estos mismos individuos a la adicción más adelante en la vida, pero esto aún no se ha determinado en estudios longitudinales.207, también puede ser importante para conferir vulnerabilidad a la adicción.
Las contribuciones genéticas a la vulnerabilidad a la adicción también son importantes. Por ejemplo, los usuarios habituales de marihuana con alelos de riesgo de genes que codifican el receptor de cannabinoides 1 (CB1) o el ácido graso amino hidrolasa 1 (FAAH; la enzima que metaboliza los cannabinoides endógenos) tuvieron una mayor reactividad relacionada con el fármaco en las áreas de PFC límbico 208. Es importante destacar que tales interacciones entre genes y entornos pueden usarse para predecir futuros comportamientos desventajosos. Por ejemplo, los aumentos de 1 año en la masa corporal de las adolescentes sanas podrían predecirse mediante la activación de la OFC lateral inducida por señales relacionadas con los alimentos, pero solo en portadoras del alelo de riesgo dopaminérgico receptor de dopamina D4 (DRD4) alelo de 7 repeticiones o el alelo DRD2 TaqIA A1 209. Estudios recientes también sugieren que las interacciones entre ciertos polimorfismos y la exposición familiar a los medicamentos, incluida la prenatal, pueden influir en el desarrollo de la OFC 210, 211. Por ejemplo, un estudio reciente mostró que el volumen de la materia gris de la OFC (mOFC) medial fue modulado por el genotipo de la monoamina oxidasa A, de modo que la variante de baja actividad de este gen hizo que la materia gris del mOFC disminuyera en individuos adictos a la cocaína 212, y esto se correlacionó con el uso de cocaína de por vida más larga.
Box 1 Cambios relacionados con la adicción en la conectividad y la estructura de PFC
La corteza prefrontal (PFC) está densamente interconectada con otras regiones y redes cerebrales corticales y subcorticales, incluida la «red de modo predeterminado» (DMN) y las «redes de atención dorsal», que están implicadas en los procesos de control ejecutivo, como la atención y la inhibición 43, 155, 156. Aunque la pregunta de cómo estas redes, y otras regiones cerebrales interconectadas, impactan en la adicción a las drogas apenas ha comenzado a ser explorada, los estudios de conectividad funcional en estado de reposo ya han demostrado ser prometedores para revelar patrones que predicen la gravedad de la enfermedad y los resultados del tratamiento. Por ejemplo, en los fumadores de cigarrillos, la corteza cingulada anterior dorsal (dACC), la conectividad estriatal se correlaciona inversamente con la gravedad de la adicción a la nicotina; el uso de un parche de nicotina mejoró significativamente la fuerza de coherencia de varias rutas de conectividad ACC, incluidas las de las estructuras de la línea media frontal 157. Además, en los fumadores abstinentes, la mejoría de los síntomas de abstinencia después de la terapia de reemplazo de nicotina se asoció con una mayor correlación inversa entre la red de control ejecutivo y la DMN, con la conectividad funcional alterada dentro de la DMN y con la conectividad funcional alterada entre la red de control ejecutivo y las regiones Implicado en la recompensa 158. Estudios más recientes sobre la adicción a la nicotina adaptaron un importante enfoque de imágenes múltiples en el que se explora la conectividad con respecto a la integridad de la materia gris y la reactividad de la señal 159, 160.
La conectividad funcional específica de la red también se reduce en otras adicciones. En individuos adictos a la cocaína, el ACC rostroventral (parte de la DMN) tuvo una conectividad más baja con el cerebro medio, donde se localizan las neuronas de dopamina 161, y se han informado resultados similares en otros estudios 162. También se han informado reducciones en la conectividad funcional en la adicción a la heroína 163, en la cual la conectividad se moduló mediante señales 164 relacionadas con las drogas y se asoció con una mayor duración del uso de heroína 165. Se necesitan más estudios para determinar si la conectividad en estado de reposo predice el desempeño de la tarea y cómo las drogas de abuso o los medicamentos potenciales cambian estas medidas; por ejemplo, ¿la administración de drogas aumenta tanto la conectividad del cerebro en reposo como las activaciones inducidas por la tarea o podría ¿Se puede asociar el estado de referencia con activaciones reducidas inducidas por tareas? Estas preguntas son importantes porque las respuestas ayudarán a determinar los puntos finales clínicos adaptados individualmente; por ejemplo, la dosis de la medicación podría reducirse según la propia conectividad funcional de estado de reposo de línea de base de un individuo.
Los estudios de imágenes estructurales han demostrado una reducción en la densidad o el grosor de la sustancia gris PFC en las poblaciones de adicciones (hasta un 20% de pérdida). Por ejemplo, los decrementos de PFC de materia gris, específicamente en el PFC dorsolateral (DLPFC), se han documentado en individuos que son adictos al alcohol. Estos decrementos se asocian con alcohol vida útil más larga usar 166, 167 y peor función ejecutiva 167, y persisten desde 6-9 meses hasta 6 años o más de abstinencia 168 – 170. A pesar de algunos resultados contradictorios 171, la mayoría de los estudios en individuos que son adictos a la cocaína 172 – 174, metanfetamina 175 , heroína 176 (incluso cuando están en terapia de reemplazo con metadona 177, 178) y la nicotina 159, 160, 179, 180 reportan reducciones similares de la sustancia gris PFC, que son más evidentes en la DLPFC, el ACC y la corteza orbitofrontal (OFC), que se asocian con una duración más prolongada o Mayor severidad del uso de drogas. La persistencia de estos cambios estructurales más allá del final del uso de drogas y hacia la abstinencia a largo plazo sugiere una influencia de factores estables o premórbidos que podrían predisponer a los individuos al uso de drogas y la adicción durante el desarrollo (RECUADRO 3). Sin embargo, tales anormalidades estructurales no se ven en los usuarios adolescentes de alcohol 181 o marihuana 182, lo que sugiere que estos decrementos de PFC también podrían ser una consecuencia dependiente de la dosis del consumo de drogas. Ya sea que predisponga a la adicción o sea una consecuencia de la adicción, tal volumen de materia gris PFC inferior, particularmente en la OFC medial, se asocia con una toma de decisiones desventajosa 183 que podría tener consecuencias catastróficas en la vida de las personas adictas.
Este estudio fue apoyado por subvenciones del Instituto Nacional de Abuso de Drogas de EE. UU. (R01DA023579 a RZG), el programa Intramural NIAAA y el Departamento de Energía, Oficina de Investigación Biológica y Ambiental (para apoyo de infraestructura). Agradecemos la contribución de AB Konova al diseño de la figura 2. Estamos en deuda con nuestros revisores cuyos comentarios fueron muy apreciados y orientaron nuestra revisión del manuscrito original.
Glosario
18Fluorodesoxiglucosa PET
( 18 F-PET). Tomografía por emisión de positrones (PET) con un radioligando para captar la captación regional de glucosa, una medida de la actividad metabólica que también se puede usar para evaluar la función cerebral global.
Metilfenidato
(MPH). Un estimulante leve (aprobado para el tratamiento del trastorno por déficit de atención con hiperactividad) con efectos farmacológicos similares a los de la cocaína (bloquea el transportador de dopamina) pero con menor potencial de abuso debido a las tasas más lentas de eliminación de la sinapsis.
Administración no contingente
Administración de un determinado medicamento que no depende del comportamiento del sujeto
Autoadministración de tasa fija.
La autoadministración de un determinado medicamento en una relación entre la administración del medicamento y el comportamiento que fija un experimentador (por ejemplo, después de la emisión de un cierto número de respuestas o después de un cierto tiempo después de la respuesta anterior).
Etiquetado de espín arterial
(También conocido como etiquetado de espín arterial). Una técnica de RM que es capaz de medir el flujo sanguíneo cerebral in vivo . Proporciona mapas de perfusión cerebral sin requerir la administración de un agente de contraste o el uso de radiación ionizante, ya que utiliza agua de sangre endógena marcada magnéticamente como un marcador de difusión libre.
Señal enmascarada
Una señal que se presenta por debajo del nivel de procesamiento consciente (es decir, fuera del conocimiento consciente). Esto generalmente se logra con una duración muy corta de la presentación de la señal seguida por la presentación de otra señal que se percibe conscientemente (mayor duración).
Ketamina
Un antagonista del receptor de NMDA utilizado principalmente para la inducción y el mantenimiento de la anestesia general. Además, puede inducir analgesia, presión arterial elevada y alucinaciones, y se ha utilizado como una droga recreativa.
[ 11 C] carfentanilo
Un radioligando receptor de tomografía por emisión de positrones (PET) que compite con opiáceos endógenos por la unión al receptor de opiáceos mu.
Afectar el emparejamiento
Una prueba neuropsicológica en la que las imágenes de caras se combinan en función de sus expresiones faciales emocionales. Esta tarea se puede utilizar para evaluar las deficiencias en el procesamiento emocional (o social).
Tarea de ir / no ir
Una tarea neuropsicológica que se usa comúnmente para evaluar el control inhibitorio. Los sujetos deben presionar un botón cuando aparece un tipo de estímulo y retener una respuesta cuando aparece otro tipo de estímulo.
Detener la señal de tiempo de reacción tarea
(SSRT). Una prueba neuropsicológica que mide la capacidad de detener una respuesta que ya se ha iniciado. Se utiliza clínicamente como índice de control inhibitorio. Un SSRT más lento está asociado con la interrupción de las funciones ejecutivas.
Errores de omisión y comisión.
Errores en una tarea de ir / no ir: un sujeto tuvo que ir pero no lo hizo (omisión de una respuesta) o tuvo que retener una respuesta pero presionó un botón en su lugar (comisión de una respuesta innecesaria). El primero es un índice de falta de atención, mientras que el segundo es un índice de respuesta impulsiva (prematura).
Tarea de Stroop
Una tarea neuropsicológica en la que se crea un conflicto entre una respuesta automática (por ejemplo, lectura) y una respuesta más lenta (por ejemplo, nombres de color), con ambos compitiendo por los mismos recursos de procesamiento. El rendimiento deficiente en las tareas de Stroop se asocia con disfunción de la corteza prefrontal.
Alexitimia
Un estado de deficiencia en la comprensión, el procesamiento o la descripción de emociones, incluida la dificultad para identificar y / o describir los propios sentimientos y el pensamiento orientado externamente.
Notas al pie
Declaración de intereses en competencia Los autores declaran no tener intereses financieros en competencia.
Volkow ND, Fowler JS. Addiction, a disease of compulsion and drive: involvement of the orbitofrontal cortex. Cereb. Cortex. 2000;10:318–325. [PubMed]
Robinson TE, Gorny G, Mitton E, Kolb B. Cocaine self-administration alters the morphology of dendrites and dendritic spines in the nucleus accumbens and neocortex. Synapse. 2001;39:257–266.[PubMed]
Robinson TE, Kolb B. Alterations in the morphology of dendrites and dendritic spines in the nucleus accumbens and prefrontal cortex following repeated treatment with amphetamine or cocaine. Eur. J. Neurosci. 1999;11:1598–1604. [PubMed]
Mansouri FA, Tanaka K, Buckley MJ. Conflict-induced behavioural adjustment: a clue to the executive functions of the prefrontal cortex. Nature Rev. Neurosci. 2009;10:141–152. [PubMed]
Kufahl PR, et al. Neural responses to acute cocaine administration in the human brain detected by fMRI. Neuroimage. 2005;28:904–914. [PubMed]
Kufahl P, et al. Expectation modulates human brain responses to acute cocaine: a functional magnetic resonance imaging study. Biol. Psychiatry. 2008;63:222–230. [PubMed]
Volkow ND, et al. Expectation enhances the regional brain metabolic and the reinforcing effects of stimulants in cocaine abusers. J. Neurosci. 2003;23:11461–11468. This study shows that the regional brain activation induced by intravenous MPH is influenced by the expectation that the subjects have when the drug is given, indicating that drug effects in an addicted individual are not just a function of the pharmacological characteristics of the drug but of past experiences and the expectations that these generate.
Zubieta JK, et al. Regional cerebral blood flow responses to smoking in tobacco smokers after overnight abstinence. Am. J. Psychiatry. 2005;162:567–577. [PubMed]
Sell LA, et al. Neural responses associated with cue evoked emotional states and heroin in opiate addicts. Drug Alcohol Depend. 2000;60:207–216. [PubMed]
Domino EF, et al. Effects of nicotine on regional cerebral glucose metabolism in awake resting tobacco smokers. Neuroscience. 2000;101:277–282. [PubMed]
Myrick H, et al. Differential brain activity in alcoholics and social drinkers to alcohol cues: relationship to craving. Neuropsychopharmacology. 2004;29:393–402. [PubMed]
de Greck M, et al. Decreased neural activity in reward circuitry during personal reference in abstinent alcoholics-a fMRI study. Hum. Brain Mapp. 2009;30:1691–1704. [PubMed]
Zijlstra F, Veltman DJ, Booij J, van den Brink W, Franken IH. Neurobiological substrates of cueelicited craving and anhedonia in recently abstinent opioid-dependent males. Drug Alcohol Depend. 2009;99:183–192. [PubMed]
Yalachkov Y, Kaiser J, Naumer MJ. Brain regions related to tool use and action knowledge reflect nicotine dependence. J. Neurosci. 2009;29:4922–4929. [PubMed]
Heinz A, et al. Brain activation elicited by affectively positive stimuli is associated with a lower risk of relapse in detoxified alcoholic subjects. Alcohol. Clin. Exp. Res. 2007;31:1138–1147. [PubMed]
Grusser SM, et al. Cue-induced activation of the striatum and medial prefrontal cortex is associated with subsequent relapse in abstinent alcoholics. Psychopharmacology. 2004;175:296–302. [PubMed]
Garavan H, et al. Cue-induced cocaine craving: neuroanatomical specificity for drug users and drug stimuli. Am. J. Psychiatry. 2000;157:1789–1798. In cocaine users, watching a cocaine-related film induced greater ACC activation than watching a sexually explicit film. This study suggests that drug-related cues in drug-addicted individuals activate similar neuroanatomical substrates as naturally evocative stimuli in healthy controls.
Brody AL, et al. Brain metabolic changes during cigarette craving. Arch. Gen. Psychiatry. 2002;59:1162–1172. [PubMed]
Artiges E, et al. Exposure to smoking cues during an emotion recognition task can modulate limbic fMRI activation in cigarette smokers. Addict. Biol. 2009;14:469–477. [PubMed]
Zhang X, et al. Masked smoking-related images modulate brain activity in smokers. Hum. Brain Mapp. 2009;30:896–907. [PubMed]
Volkow ND, et al. Activation of orbital and medial prefrontal cortex by methylphenidate in cocaine-addicted subjects but not in controls: relevance to addiction. J. Neurosci. 2005;25:3932–3939. [PubMed]
Ko CH, et al. Brain activities associated with gaming urge of online gaming addiction. J. Psychiatr. Res. 2009;43:739–747. [PubMed]
Crockford DN, Goodyear B, Edwards J, Quickfall J, el-Guebaly N. Cue-induced brain activity in pathological gamblers. Biol. Psychiatry. 2005;58:787–795. [PubMed]
Wang GJ, et al. Brain dopamine and obesity. Lancet. 2001;357:354–357. [PubMed]
Uher R, et al. Medial prefrontal cortex activity associated with symptom provocation in eating disorders. Am. J. Psychiatry. 2004;161:1238–1246. [PubMed]
Miyake Y, et al. Neural processing of negative word stimuli concerning body image in patients with eating disorders: an fMRI study. Neuroimage. 2010;50:1333–1339. [PubMed]
McBride D, Barrett SP, Kelly JT, Aw A, Dagher A. Effects of expectancy and abstinence on the neural response to smoking cues in cigarette smokers: an fMRI study. Neuropsychopharmacology. 2006;31:2728–2738. [PubMed]
Volkow ND, et al. Cognitive control of drug craving inhibits brain reward regions in cocaine abusers. Neuroimage. 2010;49:2536–2543. This study shows that when cocaine abusers try to suppress craving, this results in inhibition of limbic brain regions that is inversely associated with activation of the right inferior frontal cortex (Brodmann area 44), which is a key region for inhibitory control.
Kober H, et al. Prefrontal-striatal pathway underlies cognitive regulation of craving. Proc. Natl Acad. Sci. USA. 2010;107:14811–14816. Considering the long-term consequences of consuming cigarettes was associated with decreased craving and decreased activity in PFC regions associated with craving, and with increased activity in PFC regions associated with cognitive control. This study offers a specific cognitive– behavioural intervention to reduce cue-induced craving.
Pelchat ML, Johnson A, Chan R, Valdez J, Ragland JD. Images of desire: food-craving activation during fMRI. Neuroimage. 2004;23:1486–1493. [PubMed]
Volkow ND, Fowler JS, Wang GJ, Swanson JM. Dopamine in drug abuse and addiction: results from imaging studies and treatment implications. Mol. Psychiatry. 2004;9:557–569. [PubMed]
Koob GF, Le Moal M. Drug addiction, dysregulation of reward, and allostasis. Neuropsychopharmacology. 2001;24:97–129. [PubMed]
Solomon RL, Corbit JD. An opponent-process theory of motivation. I. Temporal dynamics of affect. Psychol. Rev. 1974;81:119–145. [PubMed]
Solomon RL, Corbit JD. An opponent-process theory of motivation. II. Cigarette addiction. J. Abnorm. Psychol. 1973;81:158–171. [PubMed]
Rolls ET. Precis of The brain and emotion. Behav. Brain Sci. 2000;23:177–191. discussion 192–233. [PubMed]
Russell M. In: Drugs and Drug Dependence. Edwards G, editor. Lexington Books; 1976. pp. 182–187.
Gold MS. In: Substance Abuse: A Comprehensive Textbook. Lowinson JH, Ruiz P, Millman RB, Langrod JG, editors. Williams & Wilkins; 1997. pp. 181–199.
Cheetham A, Allen NB, Yucel M, Lubman DI. The role of affective dysregulation in drug addiction. Clin. Psychol. Rev. 2010;30:621–634. [PubMed]
Sinha R. The role of stress in addiction relapse. Curr. Psychiatry Rep. 2007;9:388–395. [PubMed]
Aguilar de Arcos F, Verdejo-Garcia A, Peralta-Ramirez MI, Sanchez-Barrera M, Perez-Garcia M. Experience of emotions in substance abusers exposed to images containing neutral, positive, and negative affective stimuli. Drug Alcohol Depend. 2005;78:159–167. [PubMed]
Verdejo-Garcia A, Bechara A, Recknor EC, Perez-Garcia M. Executive dysfunction in substance dependent individuals during drug use and abstinence: an examination of the behavioral, cognitive, and emotional correlates of addiction. J. Int. Neuropsychol. Soc. 2006;12:405–415. [PubMed]
Goldstein RZ, et al. Is decreased prefrontal cortical sensitivity to monetary reward associated with impaired motivation and self-control in cocaine addiction? Am. J. Psychiatry. 2007;164:43–51. Sustained monetary reward was associated with a robust neuronal activation pattern in healthy control subjects but not in cocaine-addicted subjects. In addition, this study reported results that are consistent with impaired self-awareness in cocaine addiction.
Tremblay L, Schultz W. Relative reward preference in primate orbitofrontal cortex. Nature. 1999;398:704–708. [PubMed]
Elliott R, Newman JL, Longe OA, Deakin JF. Differential response patterns in the striatum and orbitofrontal cortex to financial reward in humans: a parametric functional magnetic resonance imaging study. J. Neurosci. 2003;23:303–307. [PubMed]
Breiter HC, Aharon I, Kahneman D, Dale A, Shizgal P. Functional imaging of neural responses to expectancy and experience of monetary gains and losses. Neuron. 2001;30:619–639. [PubMed]
Kringelbach ML, O’Doherty J, Rolls ET, Andrews C. Activation of the human orbitofrontal cortex to a liquid food stimulus is correlated with its subjective pleasantness. Cereb. Cortex. 2003;13:1064–1071.[PubMed]
Knutson B, Westdorp A, Kaiser E, Hommer D. FMRI visualization of brain activity during a monetary incentive delay task. Neuroimage. 2000;12:20–27. [PubMed]
O’Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Abstract reward and punishment representations in the human orbitofrontal cortex. Nature Neurosci. 2001;4:95–102. [PubMed]
Hornak J, et al. Reward-related reversal learning after surgical excisions in orbito-frontal or dorsolateral prefrontal cortex in humans. J. Cogn. Neurosci. 2004;16:463–478. [PubMed]
Kirby KN, Petry NM. Heroin and cocaine abusers have higher discount rates for delayed rewards than alcoholics or non-drug-using controls. Addiction. 2004;99:461–471. [PubMed]
Monterosso JR, et al. Frontoparietal cortical activity of methamphetamine-dependent and comparison subjects performing a delay discounting task. Hum. Brain Mapp. 2007;28:383–393. [PubMed]
Kampman KM. What’s new in the treatment of cocaine addiction? Curr. Psychiatry Rep. 2010;12:441–447. [PubMed]
Reichel CM, Bevins RA. Competition between the conditioned rewarding effects of cocaine and novelty. Behav. Neurosci. 2008;122:140–150. [PubMed]
Mattson BJ, Williams S, Rosenblatt JS, Morrell JI. Comparison of two positive reinforcing stimuli: pups and cocaine throughout the postpartum period. Behav. Neurosci. 2001;115:683–694. [PubMed]
Zombeck JA, et al. Neuroanatomical specificity of conditioned responses to cocaine versus food in mice. Physiol. Behav. 2008;93:637–650. [PubMed]
Aigner TG, Balster RL. Choice behavior in rhesus monkeys: cocaine versus food. Science. 1978;201:534–535. [PubMed]
Woolverton WL, Anderson KG. Effects of delay to reinforcement on the choice between cocaine and food in rhesus monkeys. Psychopharmacolog. 2006;186:99–106. [PubMed]
Buhler M, et al. Nicotine dependence is characterized by disordered reward processing in a network driving motivation. Biol. Psychiatry. 2010;67:745–752. Occasional smokers showed greater behavioural responses and mesocorticolimbic reactivity to stimuli predicting monetary versus cigarette rewards, whereas in dependent smokers these responses were equal for both reward types. This suggests an imbalance in the incentive salience attributed to drug-reward-predicting versus non-drug reward predicting cues in drug addiction.
Kim YT, et al. Alterations in cortical activity of male methamphetamine abusers performing an empathy task: fMRI study. Hum. Psychopharmacol. 2010;25:63–70. [PubMed]
Wang ZX, et al. Alterations in the processing of non-drug-related affective stimuli in abstinent heroin addicts. Neuroimage. 2010;49:971–976. [PubMed]
Salloum JB, et al. Blunted rostral anterior cingulate response during a simplified decoding task of negative emotional facial expressions in alcoholic patients. Alcohol. Clin. Exp. Res. 2007;31:1490–1504.[PubMed]
Asensio S, et al. Altered neural response of the appetitive emotional system in cocaine addiction: an fMRI Study. Addict. Biol. 2010;15:504–516. [PubMed]
Deroche-Gamonet V, Belin D, Piazza PV. Evidence for addiction-like behavior in the rat. Science. 2004;305:1014–1017. [PubMed]
de Ruiter MB, et al. Response perseveration and ventral prefrontal sensitivity to reward and punishment in male problem gamblers and smokers. Neuropsychopharmacology. 2009;34:1027–1038.[PubMed]
Goldstein RZ, et al. Severity of neuropsychological impairment in cocaine and alcohol addiction: association with metabolism in the prefrontal cortex. Neuropsychologia 2004;42:1447–1458 [PubMed]
Garavan H, Hester R. The role of cognitive control in cocaine dependence. Neuropsychol. Rev. 2007;17:337–345. [PubMed]
Aharonovich E, et al. Cognitive deficits predict low treatment retention in cocaine dependent patients. Drug Alcohol Depend. 2006;81:313–322. [PubMed]
Goldstein RZ, Moeller SJ, Volkow ND. In: Neuroimaging in the Addictions. Adinoff B, Stein EA, editors. Weily; 2011.
Tarter RE, et al. Neurobehavioral disinhibition in childhood predicts early age at onset of substance use disorder. Am. J. Psychiatry. 2003;160:1078–1085. [PubMed]
Kaufman JN, Ross TJ, Stein EA, Garavan H. Cingulate hypoactivity in cocaine users during a GO-NOGO task as revealed by event-related functional magnetic resonance imaging. J. Neurosci. 2003;23:7839–7843. [PubMed]
Hester R, Garavan H. Executive dysfunction in cocaine addiction: evidence for discordant frontal, cingulate, and cerebellar activity. J. Neurosci. 2004;24:11017–11022. [PubMed]
Fu LP, et al. Impaired response inhibition function in abstinent heroin dependents: an fMRI study. Neurosci. Lett. 2008;438:322–326. [PubMed]
Leland DS, Arce E, Miller DA, Paulus MP. Anterior cingulate cortex and benefit of predictive cueing on response inhibition in stimulant dependent individuals. Biol. Psychiatry. 2008;63:184–190. Informative cueing enhanced inhibitory control in a go/no-go task, and this was correlated with enhanced ACC activation in methamphetamine-addicted individuals. This study offers a specific cognitive–behavioural intervention that could be used to enhance inhibitory control in addiction.
Stroop JR. Studies of interference in serial verbal reactions. J. Exp. Psychol. 1935;18:643–662.
Leung HC, Skudlarski P, Gatenby JC, Peterson BS, Gore JC. An event-related functional MRI study of the stroop color word interference task. Cereb. Cortex. 2000;10:552–560. [PubMed]
Bench CJ, et al. Investigations of the functional anatomy of attention using the Stroop test. Neuropsychologia. 1993;31:907–922. [PubMed]
Carter CS, van Veen V. Anterior cingulate cortex and conflict detection: an update of theory and data. Cogn. Affect. Behav. Neurosci. 2007;7:367–379. [PubMed]
Eldreth DA, Matochik JA, Cadet JL, Bolla KI. Abnormal brain activity in prefrontal brain regions in abstinent marijuana users. Neuroimage. 2004;23:914–920. [PubMed]
Ersche KD, et al. Influence of compulsivity of drug abuse on dopaminergic modulation of attentional bias in stimulant dependence. Arch. Gen. Psychiatry. 2010;67:632–644. Stimulant-dependent individuals demonstrated an attentional bias for drug-related words, which was correlated with greater cue-related activation of the left prefrontal cortex; attentional bias was greater in people with highly compulsive patterns of stimulant abuse. This study also suggests that the effects of dopaminergic challenges on attentional interference and related brain activation depend on an individual’s baseline compulsivity level.
Luijten M, et al. Neurobiological substrate of smoking related attentional bias. Neuroimage. 2010;54:2374–2381. [PubMed]
Nestor L, McCabe E, Jones J, Clancy L, Garavan H. Differences in “bottom-up” and “top-down” neural activity in current and former cigarette smokers: evidence for neural substrates which may promote nicotine abstinence through increased cognitive control. Neuroimage. 56:2258–2275. [PubMed]
Khantzian EJ. The self-medication hypothesis of addictive disorders: focus on heroin and cocaine dependence. Am. J. Psychiatry. 1985;142:1259–1264. [PubMed]
Khantzian EJ. The self-medication hypothesis of substance use disorders: a reconsideration and recent applications. Harv. Rev. Psychiatry. 1997;4:231–244. [PubMed]
Langleben DD, et al. Acute effect of methadone maintenance dose on brain FMRI response to heroin-related cues. Am. J. Psychiatry. 2008;165:390–394. [PubMed]
Goldstein RZ, et al. Oral methylphenidate normalizes cingulate activity in cocaine addiction during a salient cognitive task. Proc. Natl Acad. Sci. USA. 2010;107:16667–16672. Oral MPH decreased impulsivity in a drug-relevant emotional Stroop task, and this decrease was associated with normalization of activation in the rostroventral ACC (extending to the mOFC) and dACC in cocaine-addicted individuals. These results suggest that oral MPH may have therapeutic benefits in improving cognitive–behavioural functions in cocaine-addicted individuals.
Chiu PH, Lohrenz TM, Montague PR. Smokers’ brains compute, but ignore, a fictive error signal in a sequential investment task. Nature Neurosci. 2008;11:514–520. [PubMed]
Rinn W, Desai N, Rosenblatt H, Gastfriend DR. Addiction denial and cognitive dysfunction: a preliminary investigation. J. Neuropsychiatry Clin. Neurosci. 2002;14:52–57. [PubMed]
Hester R, Nestor L, Garavan H. Impaired error awareness and anterior cingulate cortex hypoactivity in chronic cannabis users. Neuropsychopharmacology. 2009;34:2450–2458. Cannabis users showed a deficit in awareness of commission errors, and this was associated with hypoactivity in the ACC and right insula in the go/ no-go task. This study points to deficits in the role of the ACC and insula in monitoring interoceptive awareness in drug addiction.
Payer DE, Lieberman MD, London ED. Neural correlates of affect processing and aggression in methamphetamine dependence. Arch. Gen. Psychiatry. 2010;68:271–282. The ventrolateral PFC was hypoactive during affect matching in methamphetamine-dependent subjects, and this was associated with more self-reported alexithymia, pointing to a mechanism that limits emotional insight and possibly contributes to heightened aggression in addiction.
Raichle ME, Snyder AZ. A default mode of brain function: a brief history of an evolving idea. Neuroimage. 2007;37:1083–1090. discussion 1097–1089. [PubMed]
Wang W, et al. Changes in functional connectivity of ventral anterior cingulate cortex in heroin abusers. Chin. Med. J. 2010;123:1582–1588. [PubMed]
Daglish MR, et al. Functional connectivity analysis of the neural circuits of opiate craving: “more” rather than “different”? Neuroimage. 2003;20:1964–1970. [PubMed]
Yuan K, et al. Combining spatial and temporal information to explore resting-state networks changes in abstinent heroin-dependent individuals. Neurosci. Lett. 2010;475:20–24. [PubMed]
Chanraud S, et al. Brain morphometry and cognitive performance in detoxified alcohol-dependents with preserved psychosocial functioning. Neuropsychopharmacology. 2007;32:429–438. [PubMed]
Franklin TR, et al. Decreased gray matter concentration in the insular, orbitofrontal, cingulate, and temporal cortices of cocaine patients. Biol. Psychiatry. 2002;51:134–142. [PubMed]
Matochik JA, London ED, Eldreth DA, Cadet JL, Bolla KI. Frontal cortical tissue composition in abstinent cocaine abusers: a magnetic resonance imaging study. Neuroimage. 2003;19:1095–1102.[PubMed]
Sim ME, et al. Cerebellar gray matter volume correlates with duration of cocaine use in cocaine-dependent subjects. Neuropsychopharmacology. 2007;32:2229–2237 [PubMed]
Yuan Y, et al. Gray matter density negatively correlates with duration of heroin use in young lifetime heroin-dependent individuals. Brain Cogn. 2009;71:223–228. [PubMed]
Lyoo IK, et al. Prefrontal and temporal gray matter density decreases in opiate dependence. Psychopharmacology. 2006;184:139–144. [PubMed]
Liu H, et al. Frontal and cingulate gray matter volume reduction in heroin dependence: optimized voxel-based morphometry. Psychiatry Clin. Neurosci. 2009;63:563–568. [PubMed]
Brody AL, et al. Differences between smokers and nonsmokers in regional gray matter volumes and densities. Biol. Psychiatry. 2004;55:77–84. [PubMed]
Kuhn S, Schubert F, Gallinat J. Reduced thickness of medial orbitofrontal cortex in smokers. Biol. Psychiatry. 2010;68:1061–1065. [PubMed]
Volkow ND, et al. Low level of brain dopamine D2 receptors in methamphetamine abusers: association with metabolism in the orbitofrontal cortex. Am. J. Psychiatry. 2001;158:2015–2021.[PubMed]
Volkow ND, et al. Profound decreases in dopamine release in striatum in detoxified alcoholics: possible orbitofrontal involvement. J. Neurosci. 2007;27:12700–12706. [PubMed]
Fehr C, et al. Association of low striatal dopamine d2 receptor availability with nicotine dependence similar to that seen with other drugs of abuse. Am. J. Psychiatry. 2008;165:507–514. [PubMed]
Narendran R, et al. Altered prefrontal dopaminergic function in chronic recreational ketamine users. Am. J. Psychiatry. 2005;162:2352–2359. [PubMed]
Martinez D, et al. Amphetamine-induced dopamine release: markedly blunted in cocaine dependence and predictive of the choice to self-administer cocaine. Am. J. Psychiatry. 2007;164:622–629. [PubMed]
Gorelick DA, et al. Imaging brain mu-opioid receptors in abstinent cocaine users: time course and relation to cocaine craving. Biol. Psychiatry. 2005;57:1573–1582. [PubMed]
Williams TM, et al. Brain opioid receptor binding in early abstinence from alcohol dependence and relationship to craving: an [11C]diprenorphine PET study. Eur. Neuropsychopharmacol. 2009;19:740–748.[PubMed]
Kling MA, et al. Opioid receptor imaging with positron emission tomography and [18F]cyclofoxy in long-term, methadone-treated former heroin addicts. J. Pharmacol. Exp. Ther. 2000;295:1070–1076.[PubMed]
Sekine Y, et al. Brain serotonin transporter density and aggression in abstinent methamphetamine abusers. Arch. Gen. Psychiatry. 2006;63:90–100. [PubMed]
Szabo Z, et al. Positron emission tomography imaging of the serotonin transporter in subjects with a history of alcoholism. Biol. Psychiatry. 2004;55:766–771. [PubMed]
Kalivas PW. The glutamate homeostasis hypothesis of addiction. Nature Rev. Neurosci. 2009;10:561–572. [PubMed]
Laviolette SR, Grace AA. The roles of cannabinoid and dopamine receptor systems in neural emotional learning circuits: implications for schizophrenia and addiction. Cell. Mol. Life Sci. 2006;63:1597–1613. [PubMed]
Lopez-Moreno JA, Gonzalez-Cuevas G, Moreno G, Navarro M. The pharmacology of the endocannabinoid system: functional and structural interactions with other neurotransmitter systems and their repercussions in behavioral addiction. Addict. Biol. 2008;13:160–187. [PubMed]
Rao H, et al. Altered resting cerebral blood flow in adolescents with in utero cocaine exposure revealed by perfusion functional MRI. Pediatrics. 2007;120:e1245–e1254. [PubMed]
Roberts GM, Garavan H. Evidence of increased activation underlying cognitive control in ecstasy and cannabis users. Neuroimage. 2010;52:429–435. [PubMed]
Volkow ND, et al. High levels of dopamine D2 receptors in unaffected members of alcoholic families: possible protective factors. Arch. Gen. Psychiatry. 2006;63:999–1008. [PubMed]
Lotfipour S, et al. Orbitofrontal cortex and drug use during adolescence: role of prenatal exposure to maternal smoking and BDNF genotype. Arch. Gen. Psychiatry. 2009;66:1244–1252. [PubMed]
Kringelbach ML, Rolls ET. The functional neuroanatomy of the human orbitofrontal cortex: evidence from neuroimaging and neuropsychology. Prog. Neurobiol. 2004;72:341–372. [PubMed]
Ridderinkhof KR, et al. Alcohol consumption impairs detection of performance errors in mediofrontal cortex. Science. 2002;298:2209–2211. [PubMed]
Rajkowska G, Goldman-Rakic PS. Cytoarchitectonic definition of prefrontal areas in the normal human cortex: II. Variability in locations of areas 9 and 46 and relationship to the Talairach Coordinate System. Cereb. Cortex. 1995;5:323–337. [PubMed]
Autoras: Rita Z. Goldstein* and Nora D. Volkow‡ *Medical Department, Brookhaven National Laboratory, Upton, New York 11973, USA. ‡National Institute on Alcohol Abuse and Alcoholism, Bethesda, Maryland 20892, USA, and National Institute on Drug Abuse, Bethesda, Maryland 20892, USA.Correspondence to R.Z.G. vog.lnb@nietsdlogr
Objetivo: los estudios de los procesos neurobiológicos que subyacen a la adicción a las drogas se han centrado principalmente en las estructuras subcorticales límbicas. Aquí los autores evaluaron el papel de las estructuras corticales frontales en la adicción a las drogas.
Método: se propone un modelo integrado de adicción a las drogas que abarca intoxicación, atracones, abstinencia y deseo. Este modelo y los hallazgos de los estudios de neuroimagen sobre los procesos conductuales, cognitivos y emocionales que están en el centro de la adicción a las drogas se utilizaron para analizar la participación de las estructuras frontales en la adicción a las drogas.
Resultados: la corteza orbitofrontal y la circunvolución cingulada anterior, que son regiones neuroanatómicamente conectadas con estructuras límbicas, son las áreas corticales frontales más frecuentemente implicadas en la adicción a las drogas. Se activan en sujetos adictos durante la intoxicación, el deseo y los atracones, y se desactivan durante el retiro. Estas regiones también participan en funciones cognitivas y motivacionales de orden superior, como la capacidad de rastrear, actualizar y modular la prominencia de un reforzador en función del contexto y la expectativa, y la capacidad de controlar e inhibir las respuestas prepotentes.
Conclusiones: estos resultados implican que la adicción connota procesos cognitivos y emocionales regulados corticalmente, lo que resulta en la sobrevaluación de los reforzadores de la droga, la subvaloración de los reforzadores alternativos y los déficits en el control inhibitorio para las respuestas de la droga. Estos cambios en la adicción, que los autores llaman I-RISA (inhibición de la respuesta alterada y atribución de prominencia), expanden los conceptos tradicionales de la dependencia de drogas que enfatizan las respuestas reguladas límbicamente al placer y la recompensa.
La adicción es un proceso complejo de la enfermedad del cerebro que resulta de una intoxicación recurrente por drogas y está modulada por factores genéticos, de desarrollo, experienciales y ambientales. Los cambios neurobiológicos que acompañan a la adicción a las drogas no se conocen bien. Aunque hasta hace poco se creía que la adicción implicaba predominantemente procesos de recompensa mediados por circuitos límbicos (véase, por ejemplo, el síndrome de deficiencia de recompensa [ 1 ]), los resultados de estudios recientes de neuroimágenes han implicado áreas adicionales del cerebro, especialmente la corteza frontal. Aquí resumimos los hallazgos de estos estudios de neuroimagen y los incorporamos con los resultados pertinentes de los estudios preclínicos al sugerir una base para un modelo integrado de adicción a las drogas.
La mayoría de los estudios de imágenes se han concentrado en la participación de la dopamina en el proceso de la adicción a las drogas porque la capacidad de las drogas de abuso para aumentar la concentración de dopamina en el cerebro en las regiones del cerebro límbico se considera crucial por sus efectos de refuerzo ( 2 , 3 ). Sin embargo, el aumento de la dopamina per se es insuficiente para dar cuenta del proceso de adicción, ya que las drogas de abuso aumentan la dopamina tanto en personas naive como en personas adictas. De hecho, en el caso de la adicción a la cocaína, la magnitud de la dopamina inducida por drogas aumenta, y la intensidad de los autoinformes de las propiedades de refuerzo de la droga (la «alta»), parece ser menor en los sujetos adictos que en los ingenuos ( 4). Esto implica que es probable que la participación de la dopamina en la adicción a las drogas esté mediada por cambios funcionales y estructurales en los circuitos que están modulados por la dopamina, incluida la corteza frontal. En apoyo de esta sugerencia, se encuentran los resultados de varios estudios recientes de imágenes de resonancia magnética estructural / volumétrica que documentan los cambios morfológicos en el lóbulo frontal en diversas formas de adicción a las drogas. Por ejemplo, las pérdidas de volumen del lóbulo frontal se han identificado en sujetos dependientes de cocaína ( 5 , 6 ), sujetos alcohólicos ( 7 – 9 ) y sujetos dependientes de heroína ( 5).). El último estudio observó correlaciones negativas entre los volúmenes prefrontales normalizados y los años de consumo de cocaína o heroína, lo que implica un efecto acumulativo del abuso de sustancias en los volúmenes frontales. El apoyo adicional es proporcionado por los estudios realizados en ratas. Por ejemplo, recientemente se ha demostrado que la autoadministración de cocaína, pero no de alimentos, produce cambios morfológicos en dendritas y espinas dendríticas en la corteza prefrontal y el núcleo accumbens ( 10 ). La activación de la dopamina, como se observa durante la administración de anfetamina, también suprime la inhibición de la amígdala por parte de la corteza prefrontal medial, lo que posiblemente conduce a una desinhibición de las respuestas afectivas dirigidas por los sensores ( 11).). Un proceso similar puede estar ocurriendo en la adicción a las drogas humanas, en la cual se reducen los procesos prefrontales de arriba hacia abajo (ver referencia 12 ), liberando comportamientos que normalmente se mantienen bajo estrecha vigilancia y simulando reacciones similares a estrés en las que el control inhibitorio se suspende y estimula. El comportamiento conducido se acentúa.
Si la corteza frontal y sus funciones de supervisión están de hecho reguladas por la adicción a las drogas, la relevancia de los procesos motivacionales, cognitivos superiores y de autocontrol de esta aflicción no puede ser exagerada. Específicamente, proponemos que los comportamientos y los estados motivacionales asociados que están en el centro de la adicción a las drogas son, claramente, los procesos de pérdida de conductas autodirigidas / deseadas a fórmulas automáticas dirigidas por sensores y la atribución de la atención primaria a la droga de abuso en el estado. Gastos de otros estímulos gratificantes disponibles. Nuestra hipótesis es que estos estados se evocan primero en presencia de la droga de abuso o señales condicionadas a la droga, pero luego se convierten en tendencias de acción crónicas, lo que contribuye a la recaída / atracón (compulsión conductual) y la abstinencia / deseo (compulsión mental, es decir, obsesividad), respectivamente. Así, conceptualizamos la adicción a las drogas como un síndrome de inhibición de la respuesta alterada y la atribución de prominencia y lo denominamos síndrome «I-RISA» de la adicción a las drogas. El síndrome I-RISA abarca cuatro grupos de comportamientos que están interconectados en un circuito de retroalimentación positiva ( Figura 1 ) y dependen del funcionamiento de los circuitos prefrontales con respecto a la vía de recompensa subcortical.
FIGURA 1 Manifestaciones conductuales del síndrome de la adicción a las drogas (inhibición de la respuesta alterada y atribución de saliencia) I-RISA
Síndrome I-RISA de la adicción a las drogas
Intoxicación por drogas [Drug Intoxication]
El proceso de administración de drogas a corto plazo es uno que se ha asociado tradicionalmente con concentraciones de dopamina extracelulares más altas en las regiones del cerebro límbico, en particular, el núcleo accumbens ( 13 , 14 ). Sin embargo, también hay evidencia de un aumento de la concentración de dopamina en las regiones frontales ( 15 ).
Ansia de drogas [Drug Craving]
El anhelo está asociado con la respuesta aprendida que vincula la droga y su entorno a una experiencia placentera o intensamente abrumadora. Es probable que los sustratos neuroanatómicos para la consolidación de esta memoria involucren la amígdala ( 16 , 17 ) y el hipocampo ( 18 ), pero la activación del circuito tálamo-orbitofrontal y el cingulado anterior pueden ser un elemento definitorio en la experiencia real de craving ( 19 ).
Administración compulsiva de drogas [Compulsive Drug Administration]
La autoadministración compulsiva de drogas en individuos adictos ocurre incluso cuando la droga ya no se percibe como placentera y en presencia de reacciones físicas adversas a la droga ( 20 ). Este proceso de pérdida de control y atracones de medicamentos está asociado con los circuitos dopaminérgicos, serotoninérgicos y glutamatérgicos ( 21 , 22 ) y probablemente involucre la activación del circuito tálamo-orbitofrontal y el giro cingulado anterior.
Abstinencia de drogas [Drug Withdrawal]
La administración recurrente del fármaco y la posterior retirada del fármaco dan como resultado la interrupción de los circuitos conductuales que culminan en disforia, anhedonia e irritabilidad ( 23 ), lo que posiblemente contribuya a la recaída ( 24 , 25 ). Es probable que estos cambios involucren la interrupción de los circuitos corticales frontales y neurotransmisores que incluyen dopamina, serotonina y factor liberador de corticotropina ( 26 ).
Implicación de la corteza frontal
Intoxicación
Aquí revisamos los resultados de los estudios de neuroimagen que han evaluado los efectos de la administración de fármacos en medidas funcionales, como el metabolismo de la glucosa y el flujo sanguíneo cerebral (FBC). Pocos estudios han medido la actividad cerebral regional durante la intoxicación por drogas, y la mayoría de estos estudios han empleado una exposición a una sola droga. Dichos estudios han demostrado un metabolismo de la glucosa más bajo en todo el cerebro, incluida la corteza frontal, durante la intoxicación por cocaína, morfina o alcohol ( 27 – 30 ). En contraste, la intoxicación por marihuana se asocia con niveles más altos de metabolismo de la glucosa en la corteza prefrontal, la corteza orbitofrontal y el estriado en los consumidores de marihuana pero no en los que no lo hacen ( 31).). De manera similar, se ha informado de un metabolismo más rápido en la corteza prefrontal, el cingulado anterior, la corteza orbitofrontal y el estriado en los consumidores de cocaína después de la administración secuencial de metilfenidato intravenoso, que los consumidores de la cocaína reportan como similar a la cocaína intravenosa ( 19 ). Cabe señalar que la activación en la corteza orbitofrontal solo se observó en los sujetos en los que el metilfenidato indujo un deseo intenso y en la corteza prefrontal en los sujetos en los que aumentó el estado de ánimo.
Los estudios que miden los efectos de la administración de medicamentos a corto plazo en la FBC han reportado niveles más altos de FBC prefrontal durante la intoxicación con nicotina ( 32 ), marihuana ( 33 ) y alcohol ( 34 – 36 ). Además, la activación de la corteza prefrontal derecha durante la intoxicación por alcohol se asoció con la euforia ( 35 ) y durante la intoxicación por marihuana con el sentido subjetivo de la intoxicación ( 33 ). En contraste, la cocaína redujo la CBF en todo el cerebro, incluida la corteza frontal, un efecto que podría atribuirse a los efectos vasoconstrictores de la cocaína ( 37 ).
Los estudios de mapeo durante la intoxicación por drogas con imágenes de resonancia magnética funcional (fMRI, por sus siglas en inglés) para medir la respuesta dependiente del nivel de oxigenación de la sangre (BOLD, por sus siglas en inglés) han informado sobre la activación de la corteza prefrontal y el giro cingulado anterior durante la intoxicación con cocaína, un efecto que ha estado fuertemente correlacionado con Propiedades de refuerzo del fármaco ( 38 ). Los estudios de administración de nicotina también han mostrado activación en la corteza frontal y el giro cingulado anterior coincidiendo en el tiempo con las experiencias subjetivas de «rush» y «high» ( 39 ).
Las discrepancias en los patrones de activación informados podrían reflejar los efectos vasoactivos y específicos del fármaco o la diferencia en el curso temporal de los procesos medidos (estudios de metabolismo, 30 minutos; estudios de CBF-agua, 60 segundos; estudios BOLD, 3-5 segundos). Debido a que los estudios de metabolismo y CBF-agua están limitados por su pobre resolución de tiempo, el método BOLD puede ser más adecuado para evaluar la relación entre los cambios regionales y los efectos rápidos del comportamiento inducidos por los medicamentos, como «apresuramiento» y «alto». Por otro lado, el método BOLD está limitado por su sensibilidad a los cambios vasoactivos que pueden ocurrir durante la administración del fármaco.
Para resumir, la mayoría de los estudios muestran la activación en la corteza prefrontal y el giro cingulado anterior durante la intoxicación por drogas cuando se usan los métodos CBF o BOLD. Además, la activación prefrontal parece estar asociada con la percepción subjetiva de intoxicación, los efectos de refuerzo de la droga o el estado de ánimo mejorado. También es interesante que en el caso de la marihuana o el metilfenidato, la activación de las regiones frontales se observó predominantemente en los abusadores, pero no en los sujetos no abusivos. Esto sugiere que las regiones prefrontales y el cingulado anterior están involucrados en el proceso de intoxicación y que su respuesta a los medicamentos está en parte relacionada con las experiencias previas de los medicamentos.
Deseos y atracones
La administración aguda de drogas no es necesaria para la activación de la corteza frontal en individuos previamente expuestos al fármaco de elección, en los cuales, debido a la exposición previa, el deseo solo puede ser suficiente para activar los circuitos frontolímbicos. Por lo tanto, se han demostrado niveles más altos de activación cerebral (CBF, metabolismo de la glucosa o BOLD) en áreas frontolímbicas, principalmente en la corteza prefrontal y el cingulado anterior, en consumidores de cocaína expuestos a cintas de video que muestran estímulos relacionados con las drogas ( 40 – 44 ). Los autoinformes de ansia se correlacionaron significativamente con los cambios en el metabolismo de la glucosa en la corteza prefrontal dorsolateral en un estudio ( 42 ) y con la extensión espacial de la activación en la corteza prefrontal dorsolateral y el cingulado anterior en otro (43 ). En los cinco estudios, los estímulos relacionados con las drogas provocaron ansia solo en los consumidores de cocaína y no en los sujetos de comparación, lo que señala nuevamente la importancia de la experiencia previa con las drogas. El mecanismo que subyace al deseo puede implicar el recuerdo de experiencias previas emocionalmente cargadas. De hecho, el deseo se correlacionó con la activación de la amígdala en un estudio ( 42 ), y la activación de la corteza orbitofrontal se observó cuando los consumidores de cocaína recordaron y describieron su propio método de preparación de cocaína, pero no cuando describieron su árbol genealógico familiar en un estudio de nuestro estudio. Laboratorio ( 45 ) ( Figura 2 ). Además, el deseo puede implicar la anticipación de una futura recompensa de medicamentos. El papel de la corteza prefrontal dorsolateral en la anticipación de la autoadministración inmediata de fármacos fue sugerido previamente ( 42), aunque no se ha apoyado la disociación del deseo de anticipación y de ambos de la experiencia real de la droga. Sin embargo, la experiencia real del fármaco puede estar relacionada con activaciones más circunscritas que la fase de anticipación ( 42 ), en línea con la evidencia de una disociación similar de la anticipación de una experiencia sensorial (táctil) real ( 46 ).
FIGURA 2 Activación cortical orbitofrontal en abusadores activos de cocaína durante una entrevista temática de cocaína y una entrevista de tema neutral, medida por FDG PET
Otra línea de evidencia que apoya el papel de la corteza frontal en el deseo se deriva de estudios realizados poco después del último uso de drogas por parte de los sujetos. Por ejemplo, hemos demostrado un mayor metabolismo regional de la glucosa en el cerebro, incluso en el córtex orbitofrontal y el cuerpo estriado, en los consumidores de cocaína analizados durante el retiro temprano (<1 semana desde el último consumo de cocaína) que en los sujetos de comparación normales ( 47 ). Estos niveles más altos fueron proporcionales a la intensidad del deseo, de modo que cuanto mayor es el metabolismo, mayor es el deseo del fármaco. Los resultados de un estudio de nuestro laboratorio en el que el metilfenidato incrementó la corteza orbitofrontal y el metabolismo estriatal solo en los sujetos en los que aumentó el deseo ( 19).
El problema sigue siendo que el deseo de cocaína no es una medida directa del uso compulsivo de cocaína y, de hecho, su asociación con el uso de drogas y la recaída continúa siendo cuestionada ( 48 ). Demostrar la participación del córtex orbito-frontal en la autoadministración compulsiva de drogas requeriría una investigación del abusador durante el uso real en el que el suministro de drogas no está restringido. Alternativamente, un paradigma que simula un comportamiento compulsivo (como apostar cuando ya no es beneficioso) podría ofrecer una información invaluable sobre los circuitos que subyacen a la pérdida de control en la adicción.
Retirada
Las anomalías en la corteza humana asociadas con la abstinencia de la cocaína en los consumidores habituales de cocaína se documentaron en 1988 en nuestro laboratorio ( 49 ). Demostramos que los valores relativos de CBF para la corteza prefrontal y la corteza frontal lateral izquierda eran significativamente más bajos en los consumidores de cocaína que en los sujetos de comparación normales. Un estudio de seguimiento en consumidores de cocaína activos demostró diferencias en el metabolismo regional de la glucosa en el cerebro entre los consumidores de cocaína evaluados dentro de la primera semana del último consumo de cocaína y los consumidores de cocaína evaluados 2 a 4 semanas después del último consumo de cocaína ( 47). De interés, el metabolismo de la glucosa fue mayor en la corteza orbitofrontal y en el estriado en el grupo anterior que en los sujetos de comparación normales. Durante la abstinencia más prolongada (1-6 semanas desde el último uso), se encontró que el metabolismo cerebral era más bajo en los consumidores de cocaína que en los sujetos de comparación normales, un efecto que se acentuó más en la corteza frontal ( Figura 3 ) ( 50 ).
FIGURA 3 Metabolismo relativo de la glucosa inferior en la corteza prefrontal y el giro cingulado anterior de un abusador de cocaína que en un sujeto de comparación normal
Los estudios sobre los consumidores de alcohol han proporcionado evidencia similar. Por ejemplo, se documentaron anomalías del metabolismo de la glucosa (incluso en la corteza frontal) en sujetos alcohólicos por lo demás sanos, con una duración media de abstinencia de alcohol de 11 días ( 51 ). Se ha demostrado un metabolismo estriado inferior persistente en estudios de metabolismo regional después de una abstinencia del alcohol más prolongada ( 52 ). Además, los sujetos alcohólicas han demostrado una menor sensibilidad a la menor metabolismo inducida por lorazepam, una benzodiazepina que facilita la neurotransmisión de ácido γ-aminobutírico en el circuito de corteza estriado-tálamo-orbitofrontal durante los primeros (1-4 semanas) desintoxicación ( 53 ) y en el Corteza orbitofrontal durante la desintoxicación prolongada (8–11 semanas) ( 54), sugiriendo ajustes duraderos relacionados con las drogas en estas regiones del cerebro. También se documentaron anomalías persistentes después de la desintoxicación del alcohol para el cingulado anterior ( 54 ). La actividad inferior en la corteza prefrontal en sujetos alcohólicos durante la desintoxicación también se documentó en otros laboratorios que utilizaron grupos de estudio ligeramente diferentes (alcohólicos de Cloninger-tipo 2) y técnicas (tomografía computarizada de emisión de fotón único) ( 55 ). Los sujetos alcohólicos muestran menos sensibilidad en el circuito de la corteza estriado-tálamo-orbitofrontal al agonista de la serotonina m- clorofenilpiperazina, lo que demuestra la relevancia de la serotonina en estas anomalías ( 56 ). Los estudios de nuestro laboratorio también señalan la relevancia de la dopamina en la abstinencia. Primero, documentamos que en los consumidores de cocaína durante el inicio temprano (hasta 1 mes desde el último uso de cocaína) y prolongado (hasta 4 meses desde el último uso de cocaína), la abstinencia, la respuesta de dopamina estriatal o la disponibilidad de receptores fue significativamente menor ( 4 , 57 , 58 ) que en sujetos de comparación normales. También informó inferior estriatal de dopamina D 2 de unión en la heroína receptor ( 59) y abusadores de metanfetamina ( 60 ) y en sujetos alcohólicos ( 61 ) ( Figura 4 ). Además, se encontró que los niveles más bajos de receptores D 2 estriatales estaban asociados con un metabolismo más bajo en la corteza orbitofrontal y el giro cingulado anterior en sujetos adictos a la cocaína ( 58 ) ( Figura 5 ) y en la corteza orbitofrontal en los consumidores de metanfetamina ( 62 ). Finalmente, se ha demostrado un mayor metabolismo en la circunvolución cingulada anterior en respuesta al metilfenidato, que aumenta la dopamina al bloquear el transportador de dopamina ( 19 ), lo que proporciona un apoyo adicional para el papel de la activación de la dopamina inferior en el hipometabolismo frontal en la adicción a las drogas.
FIGURA 4 Unión del receptor de dopamina D2 del estriado inferior en usuarios de drogas durante la abstinencia de la cocaína, la metanfetamina y el alcohol que en sujetos de comparación normales
Para una revisión del papel del sistema de dopamina mesolímbico y mesocortical en el control inhibitorio y los procesos motivacionales en el cerebro de roedores y primates y su impacto putativo en la búsqueda de drogas, vea Jentsch y Taylor ( 63).). Aquí mencionaremos los sistemas mesolímbico y mesocortical de dopamina, que están asociados clásicamente con el refuerzo y la adicción a las drogas. El circuito de dopamina mesolímbica, que incluye el núcleo accumbens, la amígdala y el hipocampo, se ha asociado tradicionalmente con los efectos de refuerzo agudo de un fármaco y con la memoria y las respuestas condicionadas que se han relacionado con el deseo. También es probable que esté involucrado en los cambios emocionales y de motivación que se observan en los drogadictos durante el retiro. Es probable que el circuito de dopamina mesocortical, que incluye la corteza prefrontal, la corteza orbito-frontal y el cingulado anterior, esté involucrado en la experiencia consciente de intoxicación por drogas, prominencia de incentivos, expectativa / ansia de drogas y administración compulsiva de drogas. Debido a que estos circuitos funcionan en paralelo e interactúan entre sí, es probable que un determinado comportamiento implique, en mayor o menor medida, su participación conjunta. La naturaleza de estas interacciones afecta la respuesta al fármaco. Por ejemplo, la activación de circuitos de memoria (el hipocampo y la amígdala) en asociación con un contexto relacionado con el fármaco activa la corteza orbitofrontal y el cingulado anterior en espera del reforzador, que a su vez activa las células de dopamina (64 ), lo que lleva a un aumento adicional en la sensación de deseo y una posible disminución en el control inhibitorio. Note la naturaleza circular de esta interacción: la atribución de la prominencia a un estímulo dado, que es una función de la corteza orbitofrontal, depende del valor relativo de un reforzador en comparación con los reforzadores disponibles simultáneamente ( 65 ), lo que requiere el conocimiento de la fuerza de El estímulo como reforzador, función del hipocampo y amígdala. El consumo del fármaco a su vez activará más los circuitos corticales (la corteza orbitofrontal y el cingulado anterior) en proporción a la estimulación con dopamina al favorecer la respuesta del objetivo y disminuir la actividad de fondo no relacionada con el objetivo ( 66 ). La activación de estos circuitos interactivos (Figura 6) puede ser indispensable para mantener la administración compulsiva de medicamentos observada durante el atracón y al círculo vicioso de la adición de medicamentos (Figura 7).
FIGURA 6 Interacciones de los circuitos mesocorticales y mesolímbicos en la adicción a las drogas
FIGURA 7 Modelo integrador del cerebro y el comportamiento: el síndrome I-RISA (inhibición de la respuesta alterada y atribución de saliencia) de la drogadicción
Una pregunta sigue siendo por qué algunas personas se vuelven adictas y otras, en circunstancias similares, no lo hacen. Esta pregunta es uno de los temas más desafiantes en la investigación del abuso de drogas. Los mecanismos neurobiológicos que subyacen a la vulnerabilidad a la adicción a las drogas son poco conocidos y es probable que impliquen un equilibrio entre los factores que confieren vulnerabilidad y los que protegen contra ella. Sobre la base de los estudios de imagen, no se puede descartar la posibilidad de que los cambios en la actividad frontal en sujetos adictos a las drogas puedan haber anticipado su uso de drogas y podrían haber permitido la pérdida de control y la ingesta compulsiva de drogas en estos sujetos. Pero también es posible que los cambios en la actividad frontal sean secundarios al uso recurrente de drogas y que otras variables sean responsables de la vulnerabilidad de la adicción. Nosotros y otros hemos planteado la hipótesis de que la disminución de la actividad de los receptores D2 puede poner a las personas en riesgo de comportamientos adictivos como una forma de compensar temporalmente el menor número de circuitos de recompensa regulados por D2 (67). Además, hemos demostrado que la sobreexpresión de los receptores D2 en el núcleo accumbens de ratas previamente entrenadas para autoadministrarse alcohol reduce notablemente su consumo de alcohol (68). Dado que la disponibilidad del receptor D2 está asociada positivamente con la actividad frontal en el cerebro humano (revisada en la referencia 69), esto sugiere que uno de los mecanismos por los cuales los receptores D2 regulan la autoadministración de drogas, y posiblemente el potencial de adicción, es la modulación de la actividad frontal.
Mecanismos neurocognitivos
En los siguientes párrafos, resumimos los estudios de neuroimagen funcional realizados en sujetos de comparación sanos o en poblaciones sin abuso de drogas que recientemente han implicado el circuito de la corteza estriado-tálamo-orbitofrontal en los componentes de I-RISA. Específicamente, nos enfocamos en la percepción de las relaciones respuesta-refuerzo y la inhibición de la respuesta, pero también discutiremos la expectativa de recompensa y depresión. Estos cuatro componentes se pueden ver como intrincadamente relacionados con las cuatro dimensiones de nuestro modelo de adicción a las drogas ( Figura 1 ), cada uno de los cuales puede predisponer a la adicción a las drogas; así
La intoxicación por drogas se asocia con la experiencia de sus fuertes efectos de refuerzo positivos y negativos, una asociación que se fortalece a través de la autoadministración repetida y que posiblemente impide la formación de asociaciones similares; la atribución de la atención primaria a la droga ocurre a expensas de reforzadores menos poderosos.
Concebimos el deterioro en la inhibición de la respuesta como subyacente a la experiencia de recaída y atracones. Cuando la regulación de la respuesta-refuerzo se ha reducido debido a la atribución de prominencia prominente, la desinhibición de la respuesta o la respuesta impulsiva a la saliente inmediata, se esperan recompensas relacionadas con las drogas.
La expectativa de los efectos de la droga de abuso, ya sea el estado “alto” o un estado negativo más bajo, es parte integral del deseo de las drogas.
La distimia es un síntoma central de la abstinencia, posiblemente reflejando respuestas de adaptación al aumento repetido de dopamina por drogas de abuso en los circuitos de recompensa que hacen que estos últimos respondan menos a los refuerzos naturales ( 70 – 72 ). Desde el punto de vista del comportamiento, esta menor sensibilidad en los circuitos de recompensa puede representar un deterioro generalizado en la capacidad de obtener placer de los estímulos no relacionados con las drogas, lo que lleva a un estado de anhedonia, que pone a los individuos adictos a las drogas en mayor riesgo de buscar la estimulación con drogas.
Respuestas de refuerzo de respuesta y atribución de saliencia
Con respecto a la percepción de las relaciones de respuesta-refuerzo, varios estudios recientes de fMRI han monitoreado los cambios hemodinámicos del cerebro durante la realización de tareas de juego con reforzadores monetarios (consulte la referencia 73 para una revisión). Las respuestas a las ganancias y pérdidas monetarias o al ganar y perder puntos de juego se han observado en la corteza prefrontal, la corteza orbitofrontal, el cingulado anterior y el tálamo. Las activaciones de estas áreas en la adivinación, en comparación con los informes (la corteza orbitofrontal se activó exclusivamente en la tarea de adivinación en la condición más difícil, en la que la probabilidad de ser correcta se redujo a un 25% desde el 50%), apunta a un papel único del exactitud de la respuesta y mayor dependencia de la retroalimentación en condiciones de incertidumbre inherente ( 74). De hecho, un estudio reciente de resonancia magnética funcional documentó que la imprevisibilidad de una recompensa (agua o jugo) se correlacionaba con la actividad en la corteza orbitofrontal, el tálamo y el núcleo accumbens ( 75 ). Los estudios de tomografía por emisión de positrones (PET) han demostrado de manera similar las activaciones del córtex estriado-tálamo-orbitofrontal (incluidos el cingulado anterior y el córtex prefrontal dorsolateral) en asociación con el juego ( 76 , 77 ), recibiendo una retroalimentación saliente ( 78 ) o recibiendo una recompensa monetaria ( 79 ).
El procesamiento de información emocionalmente relevante y adaptativa a la conducta puede ser el núcleo de una evaluación ventajosa de las relaciones de respuesta-refuerzo. De hecho, el papel de la corteza frontal, y específicamente el cingulado anterior, en el procesamiento emocional se ha demostrado en varios estudios de PET (por ejemplo, referencias 80 – 83 ). Coherentes con estos estudios son los resultados de un estudio de resonancia magnética magnética ( 84 ) en el que el giro frontal inferior y el cingulado anterior dorsal participaron en la toma de una decisión semántica, emocionalmente cargada versus ortográfica en una tarea verbal de no ir. Otros estudios de resonancia magnética funcional han implicado la corteza orbitofrontal y el cingulado anterior en la experiencia de sensaciones agradables ( 85) y la corteza orbitofrontal en el reconocimiento de expresiones faciales emocionales temerosas, enojadas y disgustadas en comparación con expresiones neutrales ( 86 ). De interés, en un estudio PET ( 87 ), las caras enojadas pero no tristes activaron específicamente la corteza orbitofrontal, proporcionalmente con la intensidad creciente de la emoción, mientras que la corteza cingulada anterior fue coactivada por ambas expresiones. En conjunto, los resultados de estos estudios sugieren un importante papel integrador para la corteza orbitofrontal y el cingulado anterior en el análisis de la información que conlleva la importancia emotiva, evaluativa y, a largo plazo, de supervivencia para un individuo, que comprende la atribución de saliencia. , una parte integral de nuestro síndrome I-RISA de adicción a las drogas.
Inhibición de respuesta
El otro componente del síndrome I-RISA propuesto es el control del comportamiento, que se supone se rompe en los períodos de recaída y atracones de drogas. La inhibición de la respuesta ha sido relativamente bien estudiada en los paradigmas de neuroimagen. Por ejemplo, la corteza orbitofrontal, la corteza cingulada anterior y el estriado se activaron en una tarea de ir / no ir en dos estudios de IRMf ( 88 , 89 ). Una mejor inhibición de la respuesta se asoció con un mayor volumen de activación en la corteza orbitofrontal y una menor magnitud de activación en la corteza cingulada anterior, posiblemente implicando a la corteza orbitofrontal en el esfuerzo ejercido al inhibir una respuesta y la corteza cingulada anterior en la detección de errores ( 88). El apoyo adicional para el papel del cingulado anterior en la inhibición de la respuesta, incluida la competencia de respuesta y la selección, se proporciona mediante otros estudios de fMRI ( 84 , 90 – 92 ) y PET (por ejemplo, referencia 93 ) del paradigma de ir / no ir. Además, el papel del cingulado anterior en la inhibición de la respuesta se ha establecido en estudios de supresión de las tendencias de respuesta prepotentes mediante el uso del efecto Stroop (por ejemplo, referencia 94 ). Recientemente hemos proporcionado evidencia más directa del papel del circuito prefrontal en la inhibición de la respuesta en la adicción a las drogas ( 95). Examinamos la asociación entre la interferencia de Stroop y el metabolismo relativo de la glucosa en regiones cerebrales prefrontales seleccionadas en sujetos adictos a la cocaína, sujetos alcohólicos y sujetos de comparación. Los resultados revelaron que para los sujetos adictos a la cocaína y los sujetos alcohólicos, los niveles más altos de metabolismo de la corteza orbitofrontal en la línea de base se asociaron con un menor conflicto (mayor puntuación de interferencia Stroop), mientras que para los sujetos de comparación, el metabolismo de la corteza orbitofrontal más alta se asoció con un mayor conflicto ( puntuación de interferencia de Stroop más baja), lo que sugiere un cambio en el papel de la corteza orbitofrontal en función de la adicción.
Expectativa
Apoyar el papel de la corteza frontal en la expectativa es un estudio de resonancia magnética funcional que demostró distintas regiones cerebrales y diferentes características de respuesta en anticipación al dolor en comparación con la experiencia del dolor, con la primera activando más regiones anteriores (incluida la corteza frontal media anterior) que la segunda ( 46 ). La activación de la corteza orbito-frontal también se ha asociado con la expectativa en varios estudios de PET, incluida la expectativa en tareas de atención visual ( 96 ) y en tareas que implican un shock ( 97 , 98 ).
Distimia
Finalmente, se ha demostrado una asociación entre la depresión y las anomalías prefrontales en estudios de neuroimagen realizados en pacientes deprimidos, con interrupciones sugeridas de las redes frontostriatal ( 99 ) y corticolímbica ( 100 ). Los resultados de estos estudios revelaron anormalidades en reposo en los aspectos dorsolateral, ventrolateral y medial de la corteza prefrontal y el cingulado anterior, respuestas embotadas en el cingulado anterior y la corteza prefrontal medial a los desafíos conductuales y farmacológicos, y anomalías localizadas en la corteza orbitofrontal (100 ,101). También se ha informado de una menor actividad en el estriado de pacientes deprimidos en estado de reposo y en respuesta a una tarea de tiempo de reacción y retroalimentación (102, 103).
Resumen
Los estudios de imagen revisados aquí proporcionan evidencia de la participación del córtex frontal en los diversos aspectos de la adicción a las drogas, incluido el refuerzo de las respuestas a las drogas durante la intoxicación, la activación durante el deseo y la desactivación durante la abstinencia. Es probable que la participación de la corteza frontal a lo largo de estas etapas cíclicas de adicción juegue un papel importante en los cambios cognitivos y de comportamiento cognitivos que perpetúan la autoadministración de fármacos y que se destacan en el síndrome I-RISA de adición de fármacos. Las investigaciones futuras deben centrarse en la interacción entre la atribución de prominencia de la alteración y la inhibición de la respuesta y sus posibles efectos causales o predisponentes hacia el desarrollo de la adición de drogas.
Autoras: Rita Z. Goldstein, Ph.D. and Nora D. Volkow, M.D. From Brookhaven National Laboratory.Address reprint requests to Dr. Goldstein, Brookhaven National Laboratory, Medical Research, Bldg. 490, 30 Bell Ave, Upton, NY 11973; vog.lnb@nietsdlogr (e-mail).
Referencias
Blum K, Braverman ER, Holder JM, Lubar JF, Monastra VJ, Miller D, Lubar JO, Chen TJ, Comings DE: Reward deficiency syndrome: a biogenetic model for the diagnosis and treatment of impulsive, addictive, and compulsive behaviors. J Psychoactive Drugs 2000; 32(suppl i-iv):1-112
Koob GF, Caine B, Markou A, Pulvirenti L, Weiss F: Role for the mesocortical dopamine system in the motivating effects of cocaine. NIDA Res Monogr 1994; 145:1-18,
Di Chiara G: Drug addiction as dopamine-dependent associative learning disorder. Eur J Pharmacol 1999; 375:13-30, ,
Liu X, Matochik JA, Cadet JL, London ED: Smaller volume of prefrontal lobe in polysubstance abusers: a magnetic resonance imaging study. Neuropsychopharmacology 1998; 18:243-252, ,
Franklin TR, Acton PD, Maldjian JA, Gray JD, Croft JR, Dackis CA, O’Brien CP, Childress AR: Decreased gray matter concentration in the insular, orbitofrontal, cingulate, and temporal cortices of cocaine patients. Biol Psychiatry 2002; 51:134-142, ,
Jernigan TL, Butters N, DiTraglia G, Schafer K, Smith T, Irwin M, Grant I, Schuckit M, Cermak LS: Reduced cerebral grey matter observed in alcoholics using magnetic resonance imaging. Alcohol Clin Exp Res 1991; 15:418-427, ,
Jernigan TL, Schafer K, Butters N, Cermak LS: Magnetic resonance imaging of alcoholic Korsakoff patients. Neuropsychopharmacology 1991; 4:175-186,
Pfefferbaum A, Sullivan EV, Mathalon DH, Lim KO: Frontal lobe volume loss observed with magnetic resonance imaging in older chronic alcoholics. Alcohol Clin Exp Res 1997; 21:521-529, ,
Robinson TE, Gorny G, Mitton E, Kolb B: Cocaine self-administration alters the morphology of dendrites and dendritic spines in the nucleus accumbens and neocortex. Synapse 2001; 39:257-266, ,
Rosenkranz JA, Grace AA: Dopamine attenuates prefrontal cortical suppression of sensory inputs to the basolateral amygdala of rats. J Neurosci 2001; 21:4090-4103,
Miller EK, Cohen JD: An integrative theory of prefrontal cortex function. Annu Rev Neurosci 2001; 24:167-202, ,
Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ: Cocaine receptors on dopamine transporters are related to self-administration of cocaine. Science 1987; 237:1219-1223, ,
Hurd YL, Ungerstedt U: Cocaine: an in vivo microdialysis evaluation of its acute action on dopamine transmission in rat striatum. Synapse 1989; 3:48-54, ,
Goeders NE, Smith JE: Reinforcing properties of cocaine in the medial prefrontal cortex: primary action on presynaptic dopaminergic terminals. Pharmacol Biochem Behav 1986; 25:191-199, ,
Brown EE, Fibiger HC: Differential effects of excitotoxic lesions of the amygdala on cocaine-induced conditioned locomotion and conditioned place preference. Psychopharmacology (Berl) 1993; 113:123-130, ,
Meil WM, See RE: Lesions of the basolateral amygdala abolish the ability of drug associated cues to reinstate responding during withdrawal from self-administered cocaine.Behav Brain Res 1997; 87:139-148, ,
Franklin TR, Druhan JP: Expression of Fos-related antigens in the nucleus accumbens and associated regions following exposure to a cocaine-paired environment. Eur J Neurosci 2000; 12:2097-2106, ,
Fischman MW, Schuster CR, Javaid J, Hatano Y, Davis J: Acute tolerance development to the cardiovascular and subjective effects of cocaine. J Pharmacol Exp Ther 1985; 235:677-682,
Loh EA, Roberts DC: Break-points on a progressive ratio schedule reinforced by intravenous cocaine increase following depletion of forebrain serotonin. Psychopharmacology (Berl) 1990; 101:262-266, ,
Cornish JL, Duffy P, Kalivas PW: A role for nucleus accumbens glutamate transmission in the relapse to cocaine-seeking behavior. Neuroscience 1999; 93:1359-1367, ,
Willner P, Muscat R, Papp M: Chronic mild stress-induced anhedonia: a realistic animal model of depression. Neurosci Biobehav Rev 1992; 16:525-534, ,
Hodgins DC, el-Guebaly N, Armstrong S: Prospective and retrospective reports of mood states before relapse to substance use.J Consult Clin Psychol 1995; 63:400-407, ,
Johanson CE, Fischman MW: The pharmacology of cocaine related to its abuse.Pharmacol Rev 1989; 41:3-52,
Koob GF, Le Moal M: Drug addiction, dysregulation of reward, and allostasis. Neuropsychopharmacology 2001; 24:97-129, ,
Volkow ND, Hitzemann R, Wolf AP, Logan J, Fowler JS, Christman D, Dewey SL, Schlyer D, Burr G, Vitkun S, Hirschowitz J: Acute effects of ethanol on regional brain glucose metabolism and transport. Psychiatry Res 1990; 35:39-48, ,
London ED, Broussolle EPM, Links JM, Wong DF, Cascella NG, Dannals RF, Sano M, Herning R, Snyder FR, Rippetoe LR, Toung TJK, Jaffe JH, Wagner HN Jr: Morphine-induced metabolic changes in human brain: studies with positron emission tomography and [fluorine 18]fluorodeoxyglucose. Arch Gen Psychiatry 1990; 47:73-81, ,
London ED, Cascella NG, Wong DF, Phillips RL, Dannals RF, Links JM, Herning R, Grayson R, Jaffe JH, Wagner HN Jr: Cocaine-induced reduction of glucose utilization in human brain: a study using positron emission tomography and [fluorine 18]-fluorodeoxyglucose. Arch Gen Psychiatry 1990; 47:567-574, ,
de Wit H, Metz J, Wagner N, Cooper M: Behavioral and subjective effects of ethanol: relationship to cerebral metabolism using PET. Alcohol Clin Exp Res 1990; 14:482-489, ,
Volkow ND, Gillespie H, Mullani N, Tancredi L, Grant C, Valentine A, Hollister L: Brain glucose metabolism in chronic marijuana users at baseline and during marijuana intoxication. Psychiatry Res 1996; 67:29-38, ,
Nakamura H, Tanaka A, Nomoto Y, Ueno Y, Nakayama Y: Activation of fronto-limbic system in the human brain by cigarette smoking: evaluated by a CBF measurement. Keio J Med 2000; 49(suppl 1):A122-A124
Volkow ND, Mullani N, Gould L, Adler SS, Guynn RW, Overall JE, Dewey S: Effects of acute alcohol intoxication on cerebral blood flow measured with PET. Psychiatry Res 1988; 24:201-209, ,
Tiihonen J, Kuikka J, Hakola P, Paanila J, Airaksinen J, Eronen M, Hallikainen T: Acute ethanol-induced changes in cerebral blood flow. Am J Psychiatry 1994; 151:1505-1508,
Ingvar M, Ghatan PH, Wirsen-Meurling A, Risberg J, Von Heijne G, Stone-Elander S, Ingvar DH: Alcohol activates the cerebral reward system in man. J Stud Alcohol 1998; 59:258-269, ,
Breiter HC, Gollub RL, Weisskoff RM, Kennedy DN, Makris N, Berke JD, Goodman JM, Kantor HL, Gastfriend DR, Riorden JP, Mathew RT, Rosen BR, Hyman SE: Acute effects of cocaine on human brain activity and emotion.Neuron 1997; 19:591-611, ,
Stein EA, Pankiewicz J, Harsch HH, Cho J-K, Fuller SA, Hoffmann RG, Hawkins M, Rao SM, Bandettini PA, Bloom AS: Nicotine-induced limbic cortical activation in the human brain: a functional MRI study. Am J Psychiatry 1998; 155:1009-1015
Garavan H, Pankiewicz J, Bloom A, Cho J-K, Sperry L, Ross TJ, Salmeron BJ, Risinger R, Kelley D, Stein EA: Cue-induced cocaine craving: neuroanatomical specificity for drug users and drug stimuli. Am J Psychiatry 2000; 157:1789-1798
Maas LC, Lukas SE, Kaufman MJ, Weiss RD, Daniels SL, Rogers VW, Kukes TJ, Renshaw PF: Functional magnetic resonance imaging of human brain activation during cue-induced cocaine craving. Am J Psychiatry 1998; 155:124-126
Wexler BE, Gottschalk CH, Fulbright RK, Prohovnik I, Lacadie CM, Rounsaville BJ, Gore JC:Functional magnetic resonance imaging of cocaine craving. Am J Psychiatry 2001; 158:86-95
Wang G-J, Volkow ND, Fowler JS, Cervany P, Hitzemann RJ, Pappas NR, Wong CT, Felder C: Regional brain metabolic activation during craving elicited by recall of previous drug experiences.Life Sci 1999; 64:775-784
Ploghaus A, Tracey I, Gati JS, Clare S, Menon RS, Matthews PM, Rawlins JN: Dissociating pain from its anticipation in the human brain. Science 1999; 284:1979-1981
Volkow ND, Fowler JS, Wolf AP, Hitzemann R, Dewey S, Bendriem B, Alpert R, Hoff A: Changes in brain glucose metabolism in cocaine dependence and withdrawal. Am J Psychiatry 1991; 148:621-626
Tiffany ST: A cognitive model of drug urges and drug-use behavior: role of automatic and nonautomatic processes. Psychol Rev 1990; 97:147-168
Volkow ND, Mullani N, Gould KL, Adler S, Krajewski K: Cerebral blood flow in chronic cocaine users: a study with positron emission tomography. Br J Psychiatry 1988; 152:641-648
Volkow ND, Hitzemann R, Wang G-J, Fowler JS, Wolf AP, Dewey SL, Handlesman L: Long-term frontal brain metabolic changes in cocaine abusers. Synapse 1992; 11:184-190
Volkow ND, Hitzemann R, Wang G-J, Fowler JS, Burr G, Pascani K, Dewey SL, Wolf AP: Decreased brain metabolism in neurologically intact healthy alcoholics. Am J Psychiatry 1992; 149:1016-1022
Volkow ND, Wang G-J, Hitzemann R, Fowler JS, Overall JE, Burr G, Wolf AP: Recovery of brain glucose metabolism in detoxified alcoholics. Am J Psychiatry 1994; 151:178-183
Volkow ND, Wang G-J, Hitzemann R, Fowler JS, Wolf AP, Pappas N, Biegon A, Dewey SL: Decreased cerebral response to inhibitory neurotransmission in alcoholics. Am J Psychiatry 1993; 150:417-422
Volkow ND, Wang G-J, Overall JE, Hitzemann R, Fowler JS, Pappas N, Frecska E, Piscani K: Regional brain metabolic response to lorazepam in alcoholics during early and late alcohol detoxification. Alcohol Clin Exp Res 1997; 21:1278-1284
Catafau AM, Etcheberrigaray A, Perez de los Cobos J, Estorch M, Guardia J, Flotats A, Berna L, Mari C, Casas M, Carrio I: Regional cerebral blood flow changes in chronic alcoholic patients induced by naltrexone challenge during detoxification. J Nucl Med 1999; 40:19-24
Volkow ND, Fowler JS, Wolf AP, Schlyer D, Shiue C-Y, Alpert R, Dewey SL, Logan J, Bendriem B, Christman D, Hitzemann R, Henn F: Effects of chronic cocaine abuse on postsynaptic dopamine receptors. Am J Psychiatry 1990; 147:719-724
Volkow ND, Fowler JS, Wang G-J, Hitzemann R, Logan J, Schlyer D, Dewey S, Wolf AP: Decreased dopamine D2 receptor availability is associated with reduced frontal metabolism in cocaine abusers. Synapse 1993; 14:169-177
Wang GJ, Volkow ND, Fowler JS, Logan J, Abumrad NN, Hitzemann RJ, Pappas NS, Pascani K: Dopamine D2 receptor availability in opiate-dependent subjects before and after naloxone-precipitated withdrawal. Neuropsychopharmacology1997; 16:174-182
Volkow ND, Chang L, Wang G-J, Fowler JS, Leonido-Yee M, Franceschi D, Sedler MJ, Gatley SJ, Hitzemann R, Ding Y-S, Logan J, Wong C, Miller EN:Association of dopamine transporter reduction with psychomotor impairment in methamphetamine abusers. Am J Psychiatry 2001; 158:377-382
Volkow ND, Wang G-J, Fowler JS, Logan J, Hitzemann R, Ding Y-S, Pappas N, Shea C, Piscani K: Decreases in dopamine receptors but not in dopamine transporters in alcoholics.Alcohol Clin Exp Res 1996; 20:1594-1598
Volkow ND, Chang L, Wang G-J, Fowler JS, Ding Y-S, Sedler M, Logan J, Franceschi D, Gatley J, Hitzemann R, Gifford A, Wong C, Pappas N:Low level of brain dopamine D2 receptors in methamphetamine abusers: association with metabolism in the orbitofrontal cortex. Am J Psychiatry 2001; 158:2015-2021
Jentsch JD, Taylor JR: Impulsivity resulting from frontostriatal dysfunction in drug abuse: implications for the control of behavior by reward-related stimuli. Psychopharmacology (Berl) 1999; 146:373-390
Karreman M, Moghaddam B: The prefrontal cortex regulates the basal release of dopamine in the limbic striatum: an effect mediated by ventral tegmental area. J Neurochem 1996; 66:589-598
Schultz W, Tremblay L, Hollerman JR: Reward processing in primate orbitofrontal cortex and basal ganglia. Cereb Cortex2000; 10:272-284
Kiyatkin EA, Rebec GV: Dopaminergic modulation of glutamate-induced excitations of neurons in the neostriatum and nucleus accumbens of awake, unrestrained rats. J Neurophysiol 1996; 75:142-153
Volkow ND, Wang G-J, Fowler JS, Logan J, Gatley SJ, Gifford A, Hitzemann R, Ding Y-S, Pappas N: Prediction of reinforcing responses to psychostimulants in humans by brain dopamine D2 receptor levels. Am J Psychiatry 1999; 156:1440-1443
Volkow ND, Fowler JS: Addiction, a disease of compulsion and drive: involvement of the orbitofrontal cortex. Cereb Cortex2000; 10:318-325
Cassens G, Actor C, Kling M, Schildkraut JJ: Amphetamine withdrawal: effects on threshold of intracranial reinforcement.Psychopharmacology (Berl) 1981; 73:318-322
Barr AM, Phillips AG: Withdrawal following repeated exposure to d-amphetamine decreases responding for a sucrose solution as measured by a progressive ratio schedule of reinforcement. Psychopharmacology (Berl) 1999; 141:99-106
Barr AM, Fiorino DF, Phillips AG: Effects of withdrawal from an escalating dose schedule of d-amphetamine on sexual behavior in the male rat. Pharmacol Biochem Behav 1999; 64:597-604
Breiter HC, Aharon I, Kahneman D, Dale A, Shizgal P: Functional imaging of neural responses to expectancy and experience of monetary gains and losses. Neuron 2001; 30:619-639
Berns GS, McClure SM, Pagnoni G, Montague PR: Predictability modulates human brain response to reward. J Neurosci2001; 21:2793-2798
Grant SJ, Bonson KR, Contoreggi CC, London ED: Activation of the ventromedial prefrontal cortex correlates with gambling task performance: a FDG-PET study. Abstracts of the Society for Neuroscience 1999; 25(part 2):1551
Rogers RD, Owen AM, Middleton HC, Williams EJ, Pickard JD, Sahakian BJ, Robbins TW: Choosing between small, likely rewards and large, unlikely rewards activates inferior and orbital prefrontal cortex. J Neurosci 1999; 19:9029-9038
Elliott R, Frith CD, Dolan RJ: Differential neural response to positive and negative feedback in planning and guessing tasks. Neuropsychologia 1997; 35:1395-1404
Thut G, Schultz W, Roelcke U, Nienhusmeier M, Missimer J, Maguire RP, Leenders KL: Activation of the human brain by monetary reward. Neuroreport 1997; 8:1225-1228
George MS, Ketter TA, Gill DS, Haxby JV, Ungerleider LG, Herscovitch P, Post RM: Brain regions involved in recognizing facial emotion or identity: an oxygen-15 PET study. J Neuropsychiatry Clin Neurosci 1993; 5:384-394
Lane RD, Fink GR, Chau PM, Dolan RJ: Neural activation during selective attention to subjective emotional responses.Neuroreport 1997; 8:3969-3972
Pardo JV, Pardo PJ, Raichle ME: Neural correlates of self-induced dysphoria. Am J Psychiatry 1993; 150:713-719
Rainville P, Duncan GH, Price DD, Carrier B, Bushnell MC: Pain affect encoded in human anterior cingulate but not somatosensory cortex. Science 1997; 277:968-971, ,
Elliott R, Rubinsztein JS, Sahakian BJ, Dolan RJ: Selective attention to emotional stimuli in a verbal go/no-go task: an fMRI study. Neuroreport 2000; 11:1739-1744, ,
Francis S, Rolls ET, Bowtell R, McGlone F, O’Doherty J, Browning A, Clare S, Smith E: The representation of pleasant touch in the brain and its relationship with taste and olfactory areas. Neuroreport 1999; 10:453-459, ,
Carter CS, Braver TS, Barch DM, Botvinick MM, Noll D, Cohen JD: Anterior cingulate cortex, error detection, and the online monitoring of performance. Science 1998; 280:747-749
Kiehl KA, Liddle PF, Hopfinger JB: Error processing and the rostral anterior cingulate: an event-related fMRI study. Psychophysiology 2000; 37:216-223, ,
Rubia K, Overmeyer S, Taylor E, Brammer M, Williams SCR, Simmons A, Andrew C, Bullmore ET: Functional frontalisation with age: mapping neurodevelopmental trajectories with fMRI. Neurosci Biobehav Rev 2000; 24:13-19, ,
Krams M, Rushworth MFS, Deiber MP, Frackowiak RSJ, Passingham RE: The preparation, execution and suppression of copied movements in the human brain. Exp Brain Res 1998; 120:386-398, ,
Nobre AC, Coull JT, Frith CD, Mesulam MM: Orbitofrontal cortex is activated during breaches of expectation in tasks of visual attention. Nat Neurosci 1999; 2:11-12, ,
Hugdahl K, Berardi A, Thompson WL, Kosslyn SM, Macy R, Baker DP, Alpert NM, Ledoux JE: Brain mechanisms in human classical conditioning: a PET blood flow study. Neuroreport 1995; 6:1723-1728, ,
Chua P, Krams M, Toni I, Passingham R, Dolan R: A functional anatomy of anticipatory anxiety. Neuroimage 1999; 9:563-571, ,
Robbins TW, Joyce EM, Sahakian BJ: Neuropsychology and neuroimaging of affective disorders, in Handbook of Affective Disorders. Edited by Paykel ES. London, Churchill Livingston, 1992, pp 289-310
Mayberg HS, Liotti M, Brannan SK, McGinnis S, Mahurin RK, Jerabek PA, Silva JA, Tekell JL, Martin CC, Lancaster JL, Fox PT: Reciprocal limbic-cortical function and negative mood: converging PET findings in depression and normal sadness. Am J Psychiatry 1999; 156:675-682
Elliott R, Sahakian BJ, Michael A, Paykel ES, Dolan RJ: Abnormal neural response to feedback on planning and guessing tasks in patients with unipolar depression. Psychol Med 1998; 28:559-571
Teneback CC, Nahas Z, Speer AM, Molloy M, Stallings LE, Spicer KM, Risch SC, George MS: Changes in prefrontal cortex and paralimbic activity in depression following two weeks of daily left prefrontal TMS. J Neuropsychiatry Clin Neurosci1999; 11:426-435,
Hickie I, Ward P, Scott E, Haindl W, Walker B, Dixon J, Turner K: Neo-striatal rCBF correlates of psychomotor slowing in patients with major depression. Psychiatry Res 1999; 92:75-81, ,
Resumen. Introducción. Las drogas de abuso causan adicción, la cual se caracteriza por el consumo compulsivo y fuera de control de las drogas. Durante la primera fase del consumo, tienen lugar en el cerebro diversos cambios neuroquímicos y moleculares, que llevan a la dependencia y a la adicción. Desarrollo. Todas las drogas adictivas actúan a través de mecanismos neurofisiológicos comunes, principalmente basados en la activación anómala del sistema mesolímbico dopaminérgico. Aunque el cerebro del adicto trata de adaptarse a la acción de la droga, hay cambios permanentes a nivel celular y molecular que subyacen al proceso adictivo y a la aparición de síntomas de abstinencia tras el cese del consumo de la droga. Además, estos cambios neurales parece que participan en la aparición del ‘ansia de droga’ o craving tras la abstinencia, que es un deseo incontrolado de droga, principal causa de recaída y que a menudo se inicia por estímulos asociados a la droga.
Introducción
Droga es toda sustancia natural o sintética que genera adicción, es decir, la necesidad imperiosa o compulsiva de volver a consumirla para experimentar la recompensa que produce, que es sensación de placer, euforia, alivio de la tensión, etc. Por tanto, las drogas inducen adicción y refuerzan la conducta asociada a su uso. El cese en el consumo de la droga adictiva induce abstinencia, conjunto de molestias emocionales y físicas que duran unos días, y cuya intensidad depende del tipo de droga. Evitar la abstinencia es también otra causa de búsqueda y recaída en la droga (reforzador negativo).
La diferencia de los términos dependencia y adicción es confusa. En realidad, la dependencia es el conjunto de cambios neurobiológicos que subyacen al consumo de la droga y que causan la adicción. Sin embargo, la adicción sería un grado elevado de dependencia (p. ej., la cafeína induce dependencia sin ser adictiva). En la presente revisión se utilizarán los términos dependencia y adicción de modo indistinto, según la terminología DSM, que considera que son sinónimos en lo que se refiere a las drogas adictivas. Atendiendo al grado de dependencia que generan, la Organización Mundial de la Salud (OMS) clasifica las drogas adictivas según la tabla; las drogas del grupo 1 son las que inducen mayor adicción.
Las bases neurobiológicas que subyacen a la drogodependencia son motivo de intenso estudio. La razón es clara: comprender qué induce la recompensa a la droga o qué cambios crónicos generan la adicción nos permitiría actuar terapéuticamente sobre estos procesos. Las bases neurobiológicas del proceso de drogadicción se estudian en cuatro grandes fases: estado de consumo agudo, estado de consumo crónico, abstinencia aguda y abstinencia a largo plazo [19]. Para cada fase hay datos morfofuncionales, neuroquímicos y moleculares, cuya integración nos permitirán, en un futuro, una comprensión más clara de los procesos neurobiológicos que originan la drogadicción. Los estudios se han realizado y se realizan tanto en animales de laboratorio como en el ámbito clínico. Fisiológica y bioquímicamente, se utilizan numerosas técnicas tendentes a cuantificar modificaciones en la respuesta electrofisiológica, lesiones neurales selectivas, histoquímica, marcadores proteicos, registro de liberación de neurotransmisores, etc., que no se van a detallar. Conductualmente, existen diversas pruebas de laboratorio que permiten estudiar el proceso de dependencia en roedores, en sus distintas vertientes, como la autoadministración de drogas, los tests de preferencia de lugar, de sensibilización conductual, etc. La autoadministración de drogas permite evaluar el aspecto consumidor en animales, mientras que los tests de preferencia de lugar permiten abordar el aspecto incentivo. En estos últimos, se establece un condicionamiento de contexto entre un lugar –el compartimento de un enclave, generalmente– y el consumo de droga, de modo que el animal tiende a buscar dicho lugar. En los tests de sensibilización se aborda el fenómeno de la sensibilización conductual, que se cree que resulta crítico en el proceso adictivo; el test de sensibilización locomotora es el que más se utiliza – incremento progresivo de la conducta motora tras el consumo crónico de una droga–. Finalmente, el síndrome de abstinencia se evalúa tanto en sus aspectos somáticos (síntomas físicos y vegetativos) como emocionales (test de aversión de lugar). Recientemente, el empleo de ratones knockout y trasgénicos ha abierto nuevas vías en el estudio del papel de los déficit génicos y proteicos selectivos en la adicción.
Estado de consumo aguido: las drogas adictivas actúan sobre mecanismos neurofisiológicos comunes
El consumo agudo se refiere a los primeros contactos con la droga, a través de los cuales se establece el conocimiento de sus efectos placenteros y el aprendizaje tendente a su consumo. En el ser humano, mediante estudios con tomografía por emisión de positrones (PET), se sabe que el consumo agudo de psicoestimulantes, como cocaína o anfetamina, induce la activación de diversos centros nerviosos, entre los que destacan las vías mesolímbica y mesocortical, los núcleos del rafe y la amígdala (Fig. 1). En animales de experimentación se ha observado el mismo patrón de actividad tras la administración de drogas adictivas, no sólo psicoestimulantes, sino también opiáceos, nicotina o cannabinoides; se caracteriza por aumento de la expresión de cFos y de neuroaminas –principalmente dopamina (DA) o serotonina– en dichas áreas. Otras zonas, como el área gris periacueductal o el hipotálamo lateral, podrían involucrarse, pero ello no se ha demostrado todavía incuestionablemente [27].
Aunque las drogas adictivas presentan una gran diversidad molecular y actúan sobre diversos receptores y estructuras, existe un factor común a las mismas, que es la activación de la vía mesolímbica dopaminérgica, crítica en el proceso de dependencia y adicción [14,16,24]. Esta vía nace en el área tegmental ventral (ATV), y su activación durante el consumo agudo induce el incremento en la tasa de liberación de DA y una regulación al alza en los niveles de AMPc en el núcleo accumbens (NAC) y la amígdala extendida, áreas que se relacionan decisivamente con la recompensa y con el aprendizaje para el consumo [19,28]. Así, los psicoestimulantes (anfetamina y cocaína) incrementan directamente la liberación de DA en dicha vía, por medio de la inhibición del transportador de DA (ambas) o con el aumento de la exocitosis (anfetamina). Los opiáceos actúan sobre receptores opioides tipo mu e inhiben las interneuronas gabérgicas y estimulan las neuronas dopaminérgicas de la ATV. El alcohol y la nicotina activan los circuitos locales opioides de encefalinas del ATV y estimulan las neuronas del ATV. La nicotina también actúa directamente sobre receptores nicotínicos localizados en las neuronas del ATV y del NAC, y estimulan la actividad dopaminérgica mesolímbica –demostrado recientemente en humanos por Salokangas et al [23]–. Los cannabinoides actúan sobre receptores tipo CB1 localizados en las neuronas dopaminérgicas del ATV y NAC; la fenciclidina y el éxtasis aumentan la liberación de glutamato en el ATV, que, a su vez, estimula las neuronas dopaminérgicas. Los ansiolíticos benzodiacepínicos y el alcohol actúan sobre receptores tipo GABAA en el NAC y la corteza prefrontal, que, a su vez, modulan la actividad dopaminérgica (procedente del ATV) en dichas áreas. Los centros neurales que participan en el proceso adictivo se ilustran en la figura 2.
Se cree que la vía mesolímbica dopaminérgica participa, fisiológicamente, en la creación de hábitos de conducta tras estímulos reforzadores naturales (comida, bebida, sexo). La adicción sería, por tanto, una perturbación crónica de esta vía inducida por la droga, y se crearía un hábito patológico cuyo fin es el consumo de la droga (estímulo reforzador aberrante). La activación de la vía dopaminérgica mesocortical, que también nace en el ATV, ocasiona hiperactividad dopaminérgica en la corteza prefrontal, cuyo significado no está claro, pero que podría mediar en el proceso de aprendizaje y desarrollo del hábito asociado al consumo. Es evidente que el ATV desempeña un papel de primer orden en el desarrollo de la adicción y la dependencia (véase apartado siguiente).
Finalmente, los núcleos del rafe son la fuente principal de serotonina en el encéfalo, y su activación subyace al aumento en la liberación de serotonina en la fase aguda de consumo, que parece ser que participa en fenómenos de recompensa en el NAC y la corteza frontal, en paralelo a la hiperactividad dopaminérgica, principalmente tras el consumo de psicoestimulantes (cocaína, anfetamina) y éxtasis [15].
Estado de consumo crónico: existe una neuroadaptación, pero hay cambios celulares permanentes
Experimentalmente, se han investigado los efectos neurobiológicos sobre la vía mesolímbica dopaminérgica del consumo crónico de opiáceos y psicoestimulantes, y la exposición versará sobre los conocimientos acumulados tras el empleo de dichas drogas. Durante el consumo crónico tienen lugar fenómenos de neuroadaptación, que podrían representar una reacción del organismo tendente a retomar la homeostasis o equilibrio orgánico, fundamental para la vida. Sin embargo, hay cambios críticos intracelularmente y se desarrolla una sensibilización en la liberación de DA mesolímbica, hechos que resultan de gran importancia en el establecimiento de la dependencia crónica y de la abstinencia tras el cese del consumo (Fig. 3).
En animales de experimentación y estudios post mortem del cerebro de adictos a opiáceos, se ha detectado que el consumo crónico se suele acompañar de una disminución del número de receptores celulares que responden a la droga (tras su internalización) en las neuronas de la vía mesolímbica, y no hay aumento del AMPc en centros neurales como el NAC o la amígdala, al contrario de lo que sucede en la fase de consumo agudo. Estos efectos representan fenómenos de neuroadaptación. Sin embargo, los componentes intracelulares de la vía de señalización de la adenilciclasa se regulan al alza: se detecta un aumento en los niveles de proteincinasa A (PKA) y de factores de transcripción CREB (del ingles, cAMPresponseelement-binding protein), Fra1, Fra2 y delta-Fosb, tres factores de trascripción de la familia Fos [14,24]. Se cree que la mayor concentración de PKA podría deberse a un déficit en su degradación, pues el AMPc no se encuentra regulado al alza. La mayoría de los factores comentados se regulan al alza de modo transitorio, por lo que su papel en los cambios crónicos asociados a la adicción se ha cuestionado, excepto el factor delta-Fosb, que se acumula progresivamente en el cerebro con el consumo repetido de la droga.
Los mediadores comentados participan en la activación de genes nucleares, cuyos efectos no se conocen con precisión, pero se sabe que intervienen en la remodelación sináptica. Así, se expresan factores neurotróficos estimuladores y proteínas de agregación NCAM, que inducen la formación de nuevos contactos sinápticos y gap junctions. Por ejemplo, en el ATV existen factores neurotróficos estimuladores e inhibidores que mantienen un equilibrio trófico en situación normal, pero durante el desarrollo de la dependencia se detecta, en ratas, un desajuste del equilibrio a favor de factores estimuladores (véase siguiente apartado). Se ha descrito la aparición de nuevas sinapsis en el ATV y gap junctions en el NAC tras el consumo crónico de heroína en ratas. No se sabe hasta qué grado esto subyace al desarrollo de la adicción, pero debe desempeñar un papel relevante, pues el establecimiento de gap junctions es prácticamente irreversible. En el caso de drogas opiáceas, no existe internalización de receptores opiáceos mu, lo que podría relacionarse con la fuerte dependencia que ocasionan estas drogas y con la gran hiperactivación crónica de las vías de señalización intracelulares, como el excesivo recambio (turnover) de proteínas G. Este defecto en la internalización no aparece tras la estimulación con opiáceos endógenos [26], lo que indica una diferencia crítica entre opiáceos endógenos no adictivos y opiáceos adictivos de origen exógeno.
El área tegmental ventral y la adicción mediada por sensibilización
Estudios experimentales en roedores han puesto de manifiesto que la administración o exposición repetida a drogas opiáceas y psicoestimulantes da lugar a un incremento progresivo de sus efectos conductuales y de recompensa, fenómeno llamado ‘sensibilización conductual’, lo que puede observarse incluso tras años del cese del consumo de la droga [10,12,20,25]. Como se ha comentado, numerosos estudios indican que este fenómeno de sensibilización parece mediarse por el sistema mesolímbico dopaminérgico, que nace en el ATV y se dirige a áreas de control motor y límbico, como el NAC y la amígdala, regiones cerebrales que median numerosos procesos emocionales y de aprendizaje motor [11,22,27]. La administración aguda de diversas drogas, como se ha comentado, incrementa la liberación de DA en dichas áreas, y esta liberación se refuerza con el consumo crónico de la droga, lo que se denomina ‘sensibilización dopaminérgica’ [7,14,24], fenómeno paralelo a la ‘sensibilización conductual’. Esto último representa un hecho crucial de las drogas adictivas y diferencial respecto a reforzadores naturales (comida, bebida, actividad sexual), donde no hay sensibilización dopaminérgica. Por tanto, parece que la sensibilización subyace al desarrollo de la adicción a drogas y, con toda seguridad, a la abstinencia y recaída tras el cese del consumo.
Hay dos fenómenos morfoneuroquímicos básicos en el proceso de la sensibilización: 1. El proceso se inicia por cambios permanentes sinápticos en el ATV; 2. Estos cambios se median por el incremento de la actividad de glutamato en dicha área, que, a su vez, sensibiliza los receptores D1 de DA y origina una respuesta aumentada de las neuronas dopaminérgicas de esta región [2,3,9]. Por tanto, el glutamato da lugar a fenómenos de neuroplasticidad que ocasionan la sensibilización. El incremento de la actividad dopaminérgica en ATV también se pone de manifiesto por el aumento de marcadores glutamatérgicos, como los receptores NMDR1 sitos en neuronas de DA, y de la expresión de tirosinahidroxilasa (TH). Además, en el NAC, diana del ATV, se detecta un incremento en la expresión de AMPc, PKA, y delta-Fosb [17]. Actualmente, no se conocen los mediadores moleculares de dichos fenómenos, pero existen evidencias de que factores neurotróficos liberados en la ATV en respuesta al glutamato podrían participar en los cambios sinápticos permanentes, tales como el factor básico de crecimiento fibroblástico (bFGF, en inglés), el factor neurotrófico derivado de cerebro (BDNF), la neurotrofina 3 (NT3) o el factor neurotrófico de origen glial (GDNF) [8,17,21]. La acción de factores neurotróficos mediaría cambios permanentes en el circuito neuronal del ATV, como modificaciones en el tamaño neuronal, desarrollo de filamentos gliales, atrofia de neurofilamentos, etc. [17]. De hecho, en circuitos relacionados con la memoria/aprendizaje, estos cambios plásticos debidos a factores neurotróficos acompañan a tareas de aprendizaje a largo plazo. Esto supone un interesante paralelismo entre la adicción y los procesos de memoria/aprendizaje, donde subyacen cambios plásticos de circuitos neuronales [27]. La acción de los factores neurotróficos es dual: algunos parecen inducir sensibililización, y otros, la antagonizan. Así, la administración de BDNF o GDNF en el ATV ocasiona el bloqueo de la sensibilización a la morfina o cocaína en ratas. Sin embargo, la infusión de bFGF o NT3 ocasiona un aumento de la sensibilización a la morfina o cocaína [8].
Abstinencia aguda: los cambio celulares de la fase de consumo crónico originan la sintomatología al cesar el consumo de droga
Los cambios celulares antes comentados participarían también en la emergencia de los síntomas de la abstinencia tras el consumo de droga. Además, hay fenómenos de rebote morfofuncional y neuroquímicamente. Entre ellos, destaca el incremento agudo de AMPc mesolímbico y amigdalino y, en consecuencia, de los mediadores intracelulares que participan en su vía (Fig. 4). Los cambios serían responsables de la sintomatología somática (conductual y vegetativa) y emocional de la abstinencia. En el ser humano, el cuadro clínico es de mayor o menor gravedad según el tipo de droga. Así, los opiáceos inducen una abstinencia florida con anhedonia, depresión, temblor, sudación, molestias abdominales, diarrea, hipertensión, etc. El alcohol puede producir cuadros delirantes e intensa sintomatología vegetativa. Otras drogas, como la cocaína, producen cuadros de abstinencia más leves.
Respecto a las drogas opiáceas y psicoestimulantes, morfofuncional y neuroquímicamente, tanto en humanos como animales de experimentación, se ha observado una hiperactividad en áreas mesolímbicas, de modo semejante a lo que sucede en la fase de consumo, sobre todo la amígdala y amígdala extendida, así como en otras áreas como el locus ceruleus, la sustancia gris periacueductal y el hipotálamo. Los cambios en las áreas límbicas se relacionarían con la sintomatología emocional del síndrome, mientras que los otros centros participarían en la sintomatología somática (conductual y autónoma) de la abstinencia [13]. Los cambios neuroquímicos mesolímbicos parece ser que se asemejan en otras drogas que ocasionan un marcado síndrome de abstinencia, como el alcohol y los benzodiacepínicos [16].
Respecto a las drogas opiáceas, en las que se ha centrado el estudio de nuestro laboratorio, durante el síndrome agudo de abstinencia en ratas se detecta un gran aumento de DA en la corteza prefrontal, junto a una depresión acentuada de la liberación de DA en el NAC. Nuestros estudios y los de otros autores permiten postular que los cambios en la corteza prefrontal son de naturaleza dual, tanto compensatoria como agravante del síndrome. Así, el aumento de DA prefrontal disminuye la gravedad somática del síndrome (ejemplo de mecanismo homeostático), pero, de modo reactivo, disminuye la actividad dopaminérgica en el NAC, lo que es un hecho importante en el desarrollo de la disforia emocional propia del síndrome de abstinencia [18]. Otros autores han confirmado que la disforia se relaciona con hipoactividad dopaminérgica en el NAC, pero que ésta también se debe al aumento de AMPc intracelular en dicho núcleo, que ocasiona la liberación de dinorfinas que actúan sobre receptores kappa del ATV inhibiendo la actividad de las neuronas dopaminérgicas en esta región (Fig. 5). En resumen, la disforia, que es un signo cardinal de la abstinencia, parece mediarse por una disminución de la actividad dopaminérgica mesolímbica, causada tanto por la liberación de dinorfinas desde las terminales del NAC en el ATV, como por una acción inhibitoria de origen prefrontal.
Además, se detecta un aumento de la tasa de liberación de serotonina en la amígdala extendida y en la corteza cerebral. Estudios realizados en nuestro laboratorio, tras lesiones casi completas de los núcleos del rafe (fuente principal de serotonina cerebral) en ratas, indican que no se altera el síndrome de abstinencia opiáceo en su aspecto somático, pero sí se agrava el síndrome emocional y aversivo; ello indica que la serotonina ejerce una acción antidisfórica, de naturaleza homeostática, durante el síndrome de abstinencia opiáceo. Sin embargo, se incrementa casi 10 veces el efecto protector de la clonidina en el síndrome disfórico [4]. Este hecho puede ser importante como fundamento para el empleo de fármacos con posible acción beneficiosa sobre la disforia, como bloquedores selectivos de receptores de serotonina, cuyo efecto se estudia ahora. Por otra parte, se sabe que existe un importante aumento de la actividad noradrenérgica cortical y límbica durante la abstinencia, que se relaciona con la gravedad del síndrome. De hecho, el bloqueo de la hiperactividad noradrenérgica mediante fármacos como la clonidina, es capaz de atenuar significativamente la sintomatología del síndrome de abstinencia. Antes se creía que este fenómeno se debía a una hiperactividad en el locus ceruleus, fuente principal de noradrenalina del cerebro. Sin embargo, nuestros estudios han permitido demostrar que dicho núcleo no desempeña un papel relevante [5], pues su destrucción no altera el síndrome de abstinencia opiáceo. Otros autores han detectado que la actividad noradrenérgica que se origina en los núcleos noradrenérgicos espinales (A1A2) e incide principalmente en la amígdala extendida, sí es clave en la sintomatología somática del síndrome de abstinencia [6]. Finalmente, en el área gris periacueductal se detecta una hiperactividad gabérgica, que podría relacionarse con la gravedad vegetativa del síndrome, junto con el aumento de la actividad del eje hipotálamohipofisarioadrenal (HHA), que se manifiesta por el aumento de factor liberador de corticotropina (CRF) y de hormona adrenocorticotropa (ACTH) sanguínea, así como de corticoides sanguíneos.
Abstinencia a largo plazo: la recaída
Una vez que el sujeto ha superado la fase aguda de abstinencia, comienza la fase asintomática de abstinencia a largo plazo. Durante dicha fase, el sujeto puede superar definitivamente su drogadicción con apoyo médico y psicoterápico adecuado, pero tienen lugar fenómenos importantes que suelen originar la recaída de un número importante de sujetos en el consumo de la droga: el ‘ansia de droga’ (craving, en inglés) y la ‘abstinencia condicionada’. Estos fenómenos presentan un marcado carácter psicológico. Durante el ansia de droga, el sujeto presenta deseos imperiosos de volver a consumir la droga, y durante la abstinencia condicionada aparece sintomatología abstinente, a veces sin razón aparente. Ambos fenómenos se disparan, podríamos decir, en situaciones ambientales y emocionales determinadas, como estrés, recreación de lugares asociados al consumo anterior, etc. No se conocen las bases neurobiológicas de este fenómeno, pero se sabe que se relaciona con el proceso de sensibilización de la fase de consumo crónico de la droga, que, como se ha explicado, origina cambios permanentes de los circuitos mesolímbicos. Además, existe una hiperreactividad del eje HHA que puede durar años, y que explica la liberación exagerada en estos sujetos de CRF, ACTH y corticoides suprarrenales durante situaciones de estrés. Además, estas hormonas pueden agravar la sensibilización de los circuitos dopaminérgicos y facilitar la posterior aparición de ansias de droga [18].
Bibliografía
Breiter HC, Rosen BR. Functional magnetic resonance imaging of brain reward circuitry in the human. In McGinty J, ed. Advancing from the ventral striatum to the extended amygdala. New York: Annals NY Academy Sci; 1999. p. 52347.
Cador M, Bjijou Y, Stinus L. Evidence of a complete independence of the neurobiological substrates for the induction and expression of behavioral sensitization to amphetamine. Neuroscience 1995; 65: 38595.
Cador M, Bjijou Y, Cailhol S, Stinus L. Damphetamine induced behavioral sensitization: implication of a glutamatergic medial prefrontal cortex ventral tegmental area innervation. Neuroscience 1999; 94:70521.
Caillé S, Fernández Espejo E, Stinus L. Involvement of serotonin neurotransmission in opiate dependence. Behav Pharmacology 1999; 10:S14.
Caillé S, Fernández Espejo E, Reneric JP, Cador M, Koob GF, Stinus L. Total neurochemical lesion of noradrenergic neurons of the locus coeruleus does not alter either naloxone precipitated or spontaneous opiate withdrawal nor does it influence the ability of clonidine to reverse opiate withdrawal. J Pharmacol Exp Therap 1999;290: 88192.
Delfs JM, Zhu Y, Druhan JP, Aston Jones G. Noradrenaline in the ventral forebrain is critical for opiate withdrawal induced aversion. Nature 2000; 430: 4304.
Di Chiara G, Tanda G, Bassareo V, Pontieri F, Acquas E, Fenu S, et al. Drug addiction as a disorder of associative learning: role of nucleus accumbens shell/extended amygdala. In McGinty J, ed. Advancing from the ventral striatum to the extended amygdala. New York: Annals NY Academy Sci; 1999. p. 46185.
Flores C, Stewart J. Basic fibroblast growth factor as a mediator of the effects of glutamate in the development of long term sensitization to stimulant drugs: studies in the rat. Psychopharmacology 2000; 151: 15265.
Kalivas PW, Weber B. Amphetamine injection in the ventral mesencephalon sensitizes rats to peripheral amphetamine and cocaine. J Pharmacol Exp Ther 1988; 245: 1095102.
Kalivas PW, Duffy P. Time course of extracellular dopamine and behavioral sensitization to cocaine. J Neurosci 1993; 13: 26675.
Kalivas PW, Stewart J. Dopamine transmission in the initiation and expression of drug and stress induced sensitization of motor activity. Brain Res Rev 1991; 16: 22344.
Kolta MG, Shreve P, DeSouza V, Uretsky NJ. Time course of the development of the enhanced behavioral and biochemical responses to amphetamine after pretreatment with amphetamine. Neuropharmacology 1985; 24: 8239.
Koob GF, Maldonado R, Stinus L. Neural substrates of opiate withdrawal. TINS 1992; 15: 18691.
Leshner AI. Addiction is a brain disease, and it matters. Science 1997;278: 457.
Liechti ME, Vollenweider FX. Acute psychological and physiological effects of MDMA (‘Ectasy’) after haloperidol treatment in healthy humans. Eur Neuropsychopharmacol 2000; 10: 28995.
Melichar JK, Daglish MRC, Nutt DJ. Addiction and withdrawal current views. Curr Opin Pharmacol 2001; 1: 8490.
Messer CJ, Eisch AJ, Carlezon WA, Whisler K, Shen L. Role for GDNF in biochemical and behavioral adaptations to drugs of abuse. Neuron 2000; 26: 24757.
Nestler EJ. Molecular basis of long term plasticity underlying addiction. Nature Reviews Neurosci 2001; 2: 11928.
Nestler EJ, Aghajanian GK. Molecular and cellular basis of addiction. Science 1997; 278: 5863.
Piazza PV, Deminière JM, Le Moal M, Simon H. Factors that predict vulnerability to amphetamine self administration. Science 1989; 245: 15113.
Pierce RC, Pierce Bancroft AF, Prasad BM. Neurotrophin3 contributes to the initiation of behavioral sensitization to cocaine by activating the Ras/Mitogenactivated protein kinase signal transduction cascade. J Neurosci 1999; 19: 868595.
Robinson TE, Becker JB. Enduring changes in brain and behavior produced by chronic amphetamine administration: a review and evaluation of animal models of amphetamine psychosis. Brain Res Rev 1986; 396: 15798.
Salokangas RK, Vilkman H, Ilonen T, Taiminen T, Bergman J, Haaparanta M, et al. High levels of dopamine activity in the basal ganglia of cigarette smokers. Am J Psychiatry 2000; 157: 6324.
Shippenberg TS, Elmer GI. The Neurobiology of opiate reinforcement. Critical Reviews in Neurobiology 1998; 12: 267303.
Vezina P, Pierre P, Lorrain D. The effect of previous exposure to amphetamine on drug induced locomotion and self administration of a low dose of the drug. Psychopharmacology 1999; 147: 12534.
Whistler JL, Chuang HH, Chu P, Jan LY, Von Zastrow M. Functional dissociation of mu opioid receptor signaling and endocytosis: implications for the biology of opiate tolerance and addiction. Neuron 1999; 23: 73746.

 Documento completo en Scribd:
Documento completo en Scribd: